

3.4. Биосинтез углеводов
Если микроорганизмы – автотрофы, то исходным веществом для синтеза углеводов является СО2. Синтез углеводов происходит у большинства автотрофов в цикле Кальвина (восстановительный пентозофосфатный цикл), который функционирует так же, как и у растений. Цикл Кальвина – сложный путь, включающий некоторые реакции гликолиза и окислительного пентофосфатного пути. Но для цикла Кальвина характерны два специфических фермента, не участвующие в других метаболических путях, – фосфорибулокиназа, и рибулозо-1,5-дифосфаткарбо-ксилаза.
Цикл Кальвина можно разделить на три фазы: фиксация СО2 (реакция карбоксилирования), восстановление фиксированного СО2 (реакция восстановления) и регенерация акцептора СО2.
Фиксация СО2 осуществляется с участием фермента рибулозо-1,5-дифосфаткарбоксилазы, катализирующего присоединение СО2 к рибулозо-1,5-дифосфату с образованием двух молекул 3-фосфоглицериновой кислоты (3-ФГК):
СН2О Ф
СН2О
Ф
С═О
Рибулозо-1,5-дифосфаткарбоксилаза
НСОН
+ СО2 + Н2О
НСOH
СООН
+
НСОН
СООН
СН2О
Ф
НСОН
Рибулозо-1,5-дифосфат
СН2О Ф
2 молекулы 3-ФГК



В двух последующих реакциях карбоксильная группа 3-ФГК восстанавливается до альдегидной группы – восстановление фиксированного СО2. В процессе первой реакции под действием фермента 3-фосфоглицераткиназы за счет АТФ 3-ФГК превращается в 1,3-ФГК:
СН2О Ф
СН2О Ф
АТФ АДФ
НСOH
НСОН
СООН
СОО Ф


Далее 1,3-ФГК при участии глицеральдегид-3-фосфатдегидрогеназы за счет НАДН восстанавливается до 3-фосфоглицеринового альдегида (3-ФГА):

НАДН НАД+
ФН СОО Ф СНО
|
 СН2О Ф
СН2О Ф
СН2О Ф
СН2О Ф
 НСOH
НСОН
НСOH
НСОН
Часть образовавшегося 3-ФГА последовательно при участии триозофосфатизомеразы, фруктозо-1,6-дифосфатальдолазы и 1,6-фосфо-фруктозофосфатазы сначала превращается во фруктозо-6-фосфат и далее в реакциях, катализируемых ферментами глюкозофосфатизомеразой и глюкозо-6-фосфатазой, превращается в глюкозу. Однако, если бы 3-ФГА использовался только для биосинтетических процессов, то фиксация СО2 вскоре прекратилась бы из-за недостатка рибулозо-1,5-дифосфата – акцепторов СО2. Поэтому для регенерации акцептора СО2 одна молекула фруктозо-6-фосфата и три молекулы триозофосфатов (две молекулы 3-ФГА и одна молекула фосфодиоксиацетона) взаимодействуют друг с другом при участии ферментов транскетолазы, трансальдолазы (ферменты окислительного пентозофосфатного пути) и альдолазы (фермент гликолиза) с образованием трех молекул пентозофосфатов (двух молекул ксилулозо-5-фосфата и одной молекулы рибозо-5-фосфата):
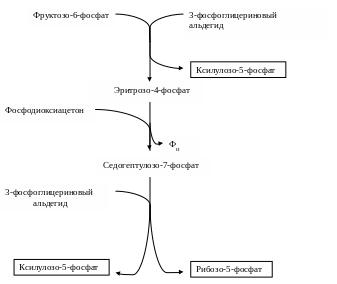
Схематически это превращение можно записать следующим образом:
С6 + 3С3 = 3С5.
Далее пентозофосфаты ксилулозо-5-фосфат и рибозо-5-фосфат превращается в рибулозо-5-фосфат под действием фосфопентозоэпимеразы и фосфопентозоизомеразы соответственно. Затем рибулозо-5-фосфат фосфорилируется вторым ферментом, участвующим только в цикле Кальвина, – фосфорибулокиназой, и таким образом регенерируется акцептор СО2 – рибулозо-1,5-дифосфат:
Фосфорибулокиназа
Р ибулозо-5-фосфат
+ АТФ Рибулозо-1,5-дифосфат
+ АДФ.
ибулозо-5-фосфат
+ АТФ Рибулозо-1,5-дифосфат
+ АДФ.
Полностью цикл Кальвина представлен на рис. 52.
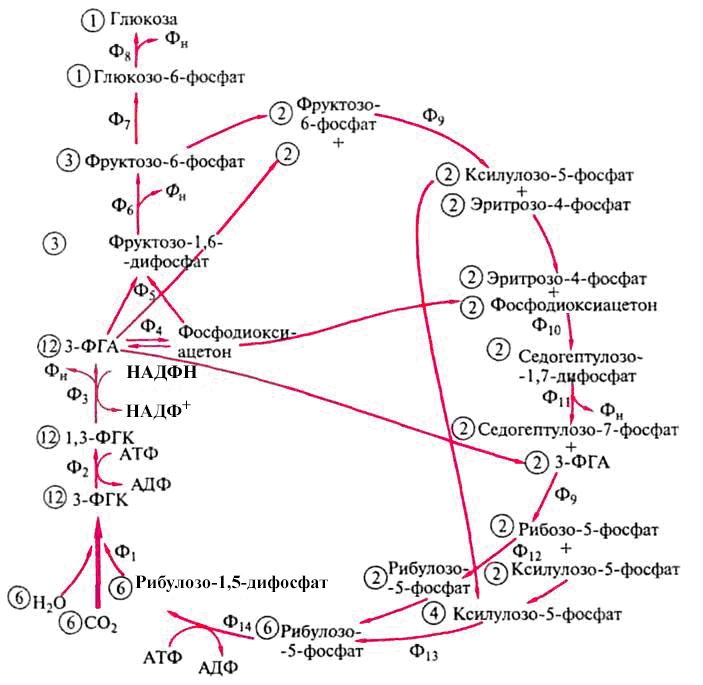
Рис. 52. Цикл Кальвина:
Ф1 – рибулозо-1,5-дифосфаткарбоксилаза; Ф2 – 3-фосфоглицераткиназа;
Ф3 – 3-ФГА-дегидрогеназа; Ф4 – триозофосфатизомераза; Ф5 – фруктозо-1,6-дифосфатальдолаза; Ф6 – 1,6-фосфофруктозофосфатаза; Ф7 – глюкозофосфатизомераза; Ф8 – глюкозо-6-фосфатаза; Ф9 – транскетолаза; Ф10 – альдолаза; Ф11 – дифосфатаза; Ф12 – фосфопентозоизомераза; Ф13 – фосфопентозоэпимераза; Ф14 – фосфорибулокиназа. Цифры, заключенные в кружок, обозначают число молекул, участвующих в реакциях (по Dagley, Nicholson, 1973)
Суммарное уравнение восстановительного пентозофосфатного цикла можно изобразить следующим образом:
6 СО2 + 18 АТФ + 12 НАДН → С6Н12О6 + 18 ФН + 18 АДФ + 12 НАД+
У бактерий-гетеротрофов на среде с неуглеводными предшественниками (например, аминокислотами, глицерином, молочной кислотой) синтез углеводов осуществляется с использованием реакций гликолитического пути, идущих в обратном направлении. Этот процесс называется глюконеогенезом. Но некоторые ферментативные реакции гликолитического пути необратимы (реакции, катализируемые гексокиназой, фосфофруктокиназой и пируваткиназой). Поэтому в клетках гетеротрофных прокариот, способных использовать двух- и трехуглеродные соединения, сформировались специальные ферментативные реакции, позволяющие обходить необратимые реакции гликолитического пути. Одной из таких обходных реакций у бактерий E. coli и других бактерий является превращение пирувата в фосфоенолпируват (ФЕП) под действием фосфоенолпируватсинтетазы:
Фосфоенолпируватсинтаза
|
 Пируват + АТФ + Н2О
ФЕП + АМФ + Фн
Пируват + АТФ + Н2О
ФЕП + АМФ + Фн
Реакция с участием фруктозодифосфатазы:
Фруктозодифосфатаза
Ф руктозо-1,6-дифосфат
+ Н2О
Фруктозо-6-фосфат + Фн.
руктозо-1,6-дифосфат
+ Н2О
Фруктозо-6-фосфат + Фн.
дает возможность обойти вторую необратимую реакцию гликолиза.
Образовавшиеся различными путями углеводы используются бактериями для синтеза олиго- и полисахаридов. Биосинтез полисахаридов осуществляется путем трансгликозилирования, т. е. путем переноса остатков моносахаридов на конец растущей цепи полисахарида. Этот процесс всегда сопровождается затратой энергии.