

Структура імунної системи організму
Центральні й периферичні органи імунітету
В даний час імунна система розглядається як система контролю, що забезпечує індивідуальність і цілісність організму. Імунна система здатна відрізнити власні структури організму від генетично чужорідних, а також переробляти і елімінувати останні.
Імунна система - це сукупність всіх лімфоїдних органів і скупчень лімфоїдних клітин, включаючи вилочкову залозу, селезінку, лімфатичні вузли, групові лімфатичні фолікули (пейерові бляшки) та інші лімфоїдні скупчення, лімфоцити кісткового мозку і периферичної крові, які складають єдиний орган імунітету (рис. 7.8).
|
Рис. 7.8. Структура імунної системи людини. |
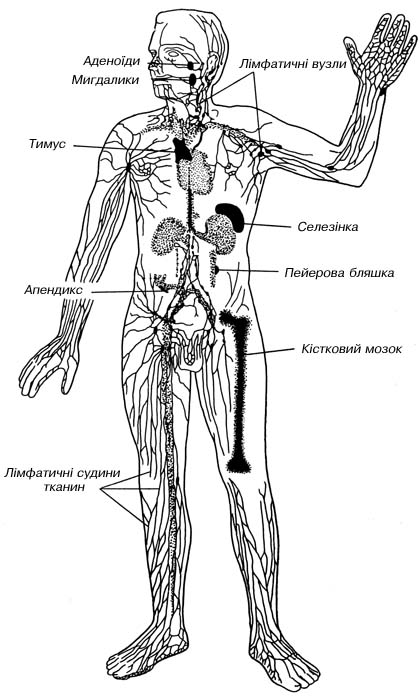
Останнім часом виділяють ще дві тканинні імунні системи - імунну систему шкіри та імунну систему слизових оболонок.
Розрізняють центральні і периферичні органи імунітету. У центральних органах, які ще називаються органами лімфопоеза, дозрівання лімфоцитів відбувається без суттєвого впливу антигенів. Розвиток периферичних органів (органів імунопоеза), навпаки, безпосередньо залежить від антигену. Лише при контакті з антигеном у них починаються процеси проліфераціі і диференціації. Центральним органом імунної системи є вилочкова залоза і сумка (бурса) Фабріціуса у птахів. У ссавців роль сумки Фабріціуса виконує кістковий мозок, який є постачальником стовбурових клітин - попередників лімфоцитів. Обидва центральних органи є місцем диференціації певних популяцій лімфоцитів. Вилоч кова залоза - джерело тимусзалежних або Т-лімфоцитів (від тимус), а в бурсі Фабріціуса (кістковому мозку) утворюються В-лімфоцити (від бурса).
Периферичними лімфоїдними органами є селезінка, лімфатичні вузли, мигдалики, лімфоїдна тканина кишечника, бронхів. На момент народження вони ще практично несформовані, тому що контакту з антигеном ще не мали. Лімфопоез здійснюється в них лише при антигенній стимуляціі. Периферичні лімфоїдні органи заселяються Т- і В-лімфоцитами із центральних органів. При цьому кожна популяція лімфоцитів мігрує в певні ділянки периферичних органів, які називаються тимусзалежними і тимуснезалежними зонами. Більшість лімфоцитів периферичних органів не залишаються в них постійно, а через деякий час, найчастіше після контакту з антигеном, включаються в рециркуляцію.
За винятком тільки деяких випадків (наприклад, передня камера ока), лімфоцити практично досягають всіх систем і органів, так що жодний антиген не залишається непоміченим.
Центральною фігурою імунної системи є лімфоцит .
Лімфоцити - це спеціалізовані клітини, які здатні реагувати (відповідати) лише на окрему групу структурно подібних антигенів. Ця здатність існує ще до першого контакту імунної системи з даним антигеном і обумовлена наявністю мембранних рецепторів, специфічних до детермінант цього антигена. Кожен клон лімфоцитів відрізняється від іншого будовою антигензв'язуючої ділянки своїх рецепторів. Таким чином, кожен клон реагує тільки на певні, відповідні йому антигени. Здатність організму відповідати практично на будь-який антиген забезпечується наявністю значної кількості різноманітних груп і клонів лімфоцитів. У результаті лімфоцити складають виключно неоднорідну популяцію клітин. Різномаїття рецепторів лімфоцитів у людини перевищує 109. Лімфоцити відрізняю ться між собою не тільки за специфічністю своїх рецепторів, але й за функціональними властивостями. За останніми розрізняють два основних класи лімфоцитів: В- лімфоцити і Т- лімфоцити (рис. 7.9).
|
Рис. 7.9. Клітини імунної відповіді: В- і Т-лімфоцити, М - макрофаги. |
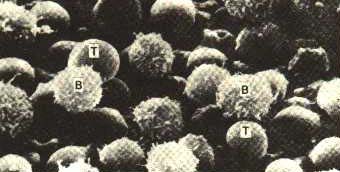
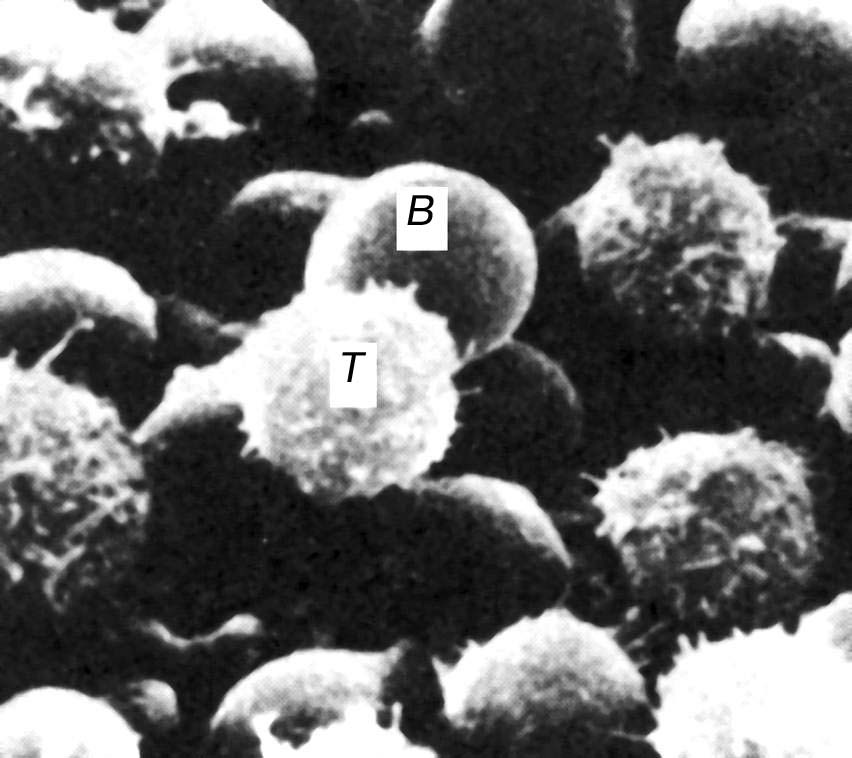

Давайте детальніше зупинимось на характеристиці цих найважливіших клітинах системи імунітету.
Характеристика В-лімфоцитів
Першою клітиною, яка започатковує появу В-лімфоцитів (рис. 7.10), є попередник В-клітини (пре-В-клітина). Найменш зрілі пре-В-клітини не синтезують ані легких, ані важких ланцюгів імуноглобу лінів, хоча вони мають антиген, спільний із зрілими В-клітинами. Ці великі пре-В-клітини діляться, утворюючи великі пре-В-клітини, що містять в цитоплазмі важкі ланцюги ІgM. Згодом, у результаті поділу останніх, виникають малі пре-В-клітини. Потім через стадію незрілих малих пре-В-клітин утворюються малі В-клітини, на мембранах яких розміщуються молекули імуноглобу лінів. Із кісткового мозку чи ембріональної печінки В-клітини потрапляють у кровообіг і мігрують у селезінку, лімфатичні вузли та інші периферичні лімфоїдні органи.
|
Рис. 7.10. В-лімфоцит на фоні формених елементів крові. |
Найменш зрілі В-клітини мають на своїй поверхні молекули ІgM. В міру дозрівання на їх поверхні з'являються IgD, рецептори компонентів комплементу, Fc - фрагментів тих імуноглобулі нів, які вони продукують. Всередині кожного клону частина В-клітин перемикається із синтезу ІgM (IgD) на синтез IgG, IgA, IgE. В-лімфоцити, в яких змінюється синтез важких ланцюгів, можуть водночас продукувати до трьох класів імуноглобулінів, наприклад: IgM, IgD, IgA.
При такій зміні ізотипів важких ланцюгів експресія генів, які визначають активний центр імуноглобуліну, не змінюється.
На поверхні зрілої В-клітини розміщуються специфічні рецептори для антигену - BcR (B cell receptor). Та ділянка рецептора, яка здатна зв'язати антиген є молекулою імуноглобуліна. Але до складу рецептора, крім імуноглобуліна, входять ще 4 поліпептид них ланцюги - по 2 з обох сторін від імуноглобуліна. Ці мембранні молекули позначаються Iga та Igb. Їх основне призначення - проведення сигналу всередину клітини про те, що антиген зв`язався з активним центром імуноглобулінового компонента. У кістковому мозку відбувається лімфопоез значної частини В-лімфоцитів (В-2 субпопуляції). Проте існує ще В-1 субпопуля ція, на клітинах якої розміщені молекули CD5 (CD - cluster definition- молекули клітинної мембрани різного призначення). На поверхні цих лімфоцитів відсутні IgD, але є IgM. Їх лімфопоез відрізняється від лімфопоезу загальновідомих В-2 лімфоцитів і полягає у тому, що попередник CD5 В-лімфоцитів ще в ембріональному періоді залишає кістковий мозок, і його нащадки у дорослих зберігаються у периферичних тканинах, в основному у черевній і плевральній порожнинах. За своєю функцією В-1 лімфоцити теж суттєво відрізняються від В-2 лімфоцитів.
В-1 лімфоцити диференціюються за межами кісткового мозку і здатні виробляти імуноглобуліни без взаємодії з Т-лімфоцитами. У більшості випадків вони продукують лише IgM. Принциповою особливістю цих імуноглобулінів є їх широка перехресна активність. Взаємодіють вони в основному з антигенами бактерій полісахаридного походження. Активація В-клітин відбувається у дві фази: проліферації і диференціації. В результаті проліферації збільшується кількість клітин, здатних реагувати із введеним в організм чужорідним антигеном. Значення проліферації велике, тому що в неімунному організмі дуже мало В-клітин, специфічних для будь-якого антигену.
Таким чином, в результаті проліферації збільшується кількість клітин, здатних негайно диференціюватись в антитілоутво рюючі клітини. При активації частина клону В-лімфоцитів знову перетворюється у малі лімфоцити і не диференцюється у плазматичні клітини. Це відносно тривалоіснуючі клітини, які значно легше активуються при повторній стимуляції, ніж вихідні В-клітини. Тому ці клітини отримали назву В-клітин пам'яті. В активованих В-лімфоцитів поступово зникають поверхневі імуноглобуліни і замість них клітини починають синтезувати молекули антитіл, що секретуються назовні. Як відомо, плазматич ні клітини відрізняються від В-лімфоцитів ексцентрично розміщеним ядром, добре розвинутим апаратом Гольджі. У цих клітин ендоплазматичний ретикулум займає майже всю цитоплазму, що необхідно для активного синтезу і продукції антитіл. Зрілі плазматичні клітини здатні синтезувати декілька тисяч молекул імуноглобулінів протягом секунди. Але така продуктивність поєднується з короткою тривалістю життя (2-3 дні).
Фази проліферації і диференціації В-лімфоцитів перебувають під впливом різноманітних факторів, серед яких найбільше значення мають інтерлейкіни (ІЛ). Більшість вчених вважає, що за активацію В-клітин відповідає ІЛ-4 , за проліферацію - ІЛ-5, диференціацію - ІЛ-6. Хоч на окремих етапах, крім цих трьох інтерлейкінів, можуть брати участь і інші цитокіни. Такі як ІЛ-1, ІЛ-2, інтерферон та інші.
Функціональна характеристика Т-лімфоцитів
Частина попередників Т-лімфоцитів із кісткового мозку та ембріональної печінки мігрують у тимус і зазнають серії перетворень у процесі диференціації. Ці попередники ще не мають характерних для Т-лімфоцитів молекул СD4, СD8, і тому їх називають подвійними негативними клітинами. Потім вони розвиваються у подвійні позитивні клітини CD4+CD8+ з наступною диференціацією у CD4+CD8- і CD4-CD8+ дозрілі клітини. У процесі розвитку спостерігається позитивна селекція з виживанням тимоцитів, які взаємодіють з власними антигенами головного комплексу гістосумісності (ГКГ), і негативна селекція з елімінацією тих клітин (аутореактивних) , які реагують із власними антигенними структурами тканин організму.
Друга частина стовбурових клітин - попередників Т-лімфоцитів попадає із кісткового мозку у слизову оболонку кишкового тракту і там диференціюється у Т-лімфоцити. Вони локалізують ся у бар'єрних тканинах, в основному у слизовій кишечника, де і виконують свої функції. Ці клітини за своїми рецепторами відрізняються від тимусної популяції Т-лімфоцитів. Їм властивий рецептор TcRgd (T cell receptor gd).
Лімфоцити - єдині клітини крові, які проходять двоетапну диференціацію. Перший етап - лімфопоез - здійснюється у кістковому мозку і тимусі. Другий етап - імуногенез є продовженням диференціації лімфоцита після його контакту з антигеном. Кожен лімфоцит здатний розпізнавати і зв`язувати тільки свій антиген.
Лімфопоез - антигеннезалежний процес. Імунопоез - антигензалежний процес, продовження диференціації. Слід пам'ятати, що синтез на мембрані антигенрозпізнавального рецептора лімфоцита відбувається під час лімфопоеза, коли ще лімфоцит не зустрічав ся з антигеном. У результаті лімфопоеза, який триває все життя, здоровий організм формує більше 109 варіантів клонів лімфоцитів. Кожен клон лімфоцитів має на своїй поверхні єдиний варіант антигензв 'язуючого рецептора (кожен лімфоцит існує для одного, свого антигена). Такою властивістю не володіють жодні клітини. Таким чином, в органи лімфопоезу попадають недиференційо вані клітини попередники. Із них виникають зрілі неімунні лімфоцити, готові зустріти антиген. Стабільним маркером неімунних Т-лімфоцитів є молекули CD45RA (повна ізоформа мембранного фермента тирозинфосфатази). У тимусі у попередників Т-лімфоцитів появляються молекули CD4 i CD8 і згодом рецептор TcRab.
Таким чином, Т-лімфоцити (рис. 7.11) можуть мати два молекулярних варіанти рецепторів для антигена - TcRab i TcRgd, і на кожному з них знаходиться лише одна різновидність рецептора: або TcRab, або TcRgd.
|
Рис. 7.11. Т-лімфоцит. |
Всі Т-лімфоцити з TcRab диференціюються із стовбурової клітини у тимусі. Більшість лімфоцитів з TcRgd диференціюю ться поза тимусом, головним чином у слизовій шлунково-киш кового тракту, і там виконують свою функцію. У крові циркулює незначна кількість Тgd-лімфоцитів, але в організмі вони складають майже половину всіх Т-лімфоцитів і локалізовані у бар'єрних органах, більшість - у слизовій шлунково-кишкового тракту.
Тимоцити з готовими рецепторами (TcR) взаємодіють з антигенами ГКГ першого або другого класів і проходять процеси позитивної і негативної селекції. Негативна селекція полягає у загибелі (за механізмом апоптозу) тимоцитів, які не зв'язали своїм рецептором антигени ГКГ, а також тимоцитів , які досить міцно зв'язали ці антигени.
Клітини моноцитарно-макрофагального ряду і інші клітини, що складають мікрооточення тимусу (клітини-няньки, епітеліальні і дендритні клітини), беруть безпосередню участь в процесі дозрівання Т-лімфоцитів. Макрофаги, напевно, одна із основних субпопуляцій клітин тимусу, що продукують метаболіти арахідонової кислоти (МАК). Синтез МАК починається на ранніх стадіях онтогенезу і регулюється гормонами тимусу (наприклад, тимуліном). Встановлено, що простагландини (продукти метаболіз му арахідонової кислоти) викликають апоптоз тимоцитів (запрограмована загибель клітин) шляхом активації ендонуклеаз і, таким чином, беруть участь в елімінації заборонених клонів лімфоцитів. З іншого боку, МАК, які утворюються під дією ліпоксиге наз (лейкотриєни), пригнічують апоптоз.
Позитивна селекція полягає у вибірковому захисті від апоптозу тимоцитів, які зв'язали який-небудь пептидний антиген із середньою афінністю. У залежності від того, яка антигенпрезентуюча клітина працює з тимоцитом на даному етапі лімфопоеза, на клітинній мембрані та у геномі тимоцита закріплюється експресія лише однієї із ключових молекул - або CD4 (яка комплементарно зв`язується з власними антигенами ГКГ ІІ класу), або CD8 (яка комплементарно зв`язується з власними антигенами ГКГ І класу). Відповідно з CD4+ тимоцита, в майбутньому у периферичних органах виникне який-небудь із Т хелперів. Із CD8+ тимоцитів утворяться цитотоксичні Т-лімфоцити ефектори. Тільки такі тимоцити у нормі будуть мігрувати з тимусу у периферичні тканини і чекати там зустрічі із своїм антигеном. Після зустрічі з анти геном у периферичних тканинах розпочинається другий етап диференціації Т-лімфоцита - імуногенез.
Серед Т-лімфоцитів розрізняють 8 великих функціонально різних субпопуляцій. Проте всі вони містять на клітинній мембрані молекули CD3, які складаються із трьох поліпептидних ланцюгів (e,g,d). Ці ланцюги закріплюють TcR по боках. З трансмембранною частиною рецептора пов'язані ще два ланцюги, які занурені всередину клітини й забезпечують передачу сигналу про те, що рецептор зв'язав антиген. Після цього запускаються відповідні реакції в ядрі і цитоплазмі.
В процесі дозрівання і диференціації появляються різні види Т-лімфоцитів, які відрізняються за функціональним призначен ням, антигенною структурою та інш.
Так розрізняють Т-лімфоцити помічники (хелпери), в основному з молекулами CD4+CD8-, лімфоцити із супресорними властвостями, Т-лімфоцити ефектори (кілери) з молекулами CD4-CD8+, Т-контрсупресори. Якщо молекули CD4 розташовані на Т-лімфоцитах з рецептором TcRab, то ці клітини називаються Т-хелперами (Th). Крім цих лімфоцитів, така молекула властива нейронам, макрофагам, еозинофілам. Недавно встановлено, що серед Th з фенотипом CD4+CD8- можна розрізнити декілька субпопуляцій, які у процесі імунної відповіді продукують різні цитокіни і позначаються Th0, Th1, Th2, Th3.
Th0 - містять гени, які характерні для всіх інших типів Т-хелперів. Тh1 - єдині Т-хелпери, які продукують g- інтерферон, інтерлейкін-2 (ІЛ-2), фактор некрозу пухлин, лімфотоксин. Вони стимулюють проліферацію Т- і В-лімфоцитів, сприяють перемикан ню синтезу імуноглобулінів у В-лімфоцитах з класу IgM на клас IgG, підсилюють активність макрофагів (g інтерферон). Th1 - основні патогномонічні клітини, що запускають реакції гіперчутли вості сповільненого типу (ГСТ). Остання якраз і розвивається у такій послідовності антиген - Th1- g інтерферон - активований макрофаг - макрофаг - запалення.
Th2 - відрізняються від Th1 за тією ознакою, що продукують ІЛ-4, ІЛ-5, ІЛ-10, ІЛ-13. Цими цитокінами обумовлені основні функціональні властивості Th2: ІЛ-4- сприяє перемиканню біосинтезу імуноглобулінів у В-лімфоцитах з класу IgM на IgE; ІЛ-5 - сильний активатор для еозинофілів - спільно з ІЛ-4 організовує захисну реакцію проти паразитів (гельмінтів, найпростіших). Th3 - ще нечітко виділена субпопуляція. До неї належать ті Т-хелпери, які у значній кількості секретують ІЛ-4, ІЛ-10, TGFb (трансформуючий фактор росту b). Цей фактор є головним гуморальним чинником супресії імунної відповіді. Клоновані клітини Th3 здатні викликати стан толерантності. Функціональна здатність Th3 відповідає поняттю Т-лімфоцитів супресорів , якими до недавна вважали Т-лімфоцити з фенотипом CD8+. У даний час клітини - супресори перестали сприймати як одну з особливих субпопуляцій Т-лімфоцитів, що розвиваються у процесі лімфопоеза. Супресія імунної відповіді вцілому - багатофакторне явище. Вона спричинена елімінацією антигена як ініціатора імунної відповіді, апоптозом простимульованих лімфоцитів, антипроліфе ративною дією трансформуючого фактора росту b, невелику кількість якого виробляють різні лімфоцити і дуже багато Th3. Але оскільки ці ж Th3 виділяють і інші цитокіни (ІЛ-4, ІЛ-10), що не мають імуносупресорного призначення, то виділяти супресорні клітини як окремої субпопуляції є не зовсім доцільним. Від функціонального стану супресорних механізмів залежить розвиток аутоімунних, імунодефіцитних, алергічних станів, пухлинних процесів, вираженість трансплантаційних реакцій, гіперчутливості сповільненого типу, розвиток імунної відповіді на тимусзалежні і тимуснезалежні антигени. Наприклад, при їх недостатності ефекторні клітини імунної системи (Т-ефектори, В-лімфоцити) отримують необмежену можливість реагувати проти власних клітин і тканин, що обумовлює появу аутоімунних і алергічних реакцій. У той же час, якщо кількісні і функціональні показники їх надто високі, то створюється сприятливий фон для розвитку імунодефіцитних захворювань. Один із гормонів вилочкової залози - тимопоетин вибірково стимулює функцію супресорних механізмів, що відіграють певну роль у регуляції цих процесів. Т-хелпери будь-якої функціональної спеціалізації об'єднує одна загальна властивість - особливість їх антигензв'язуючого рецептора TcR. TcRab здатний разом з мембранною молекулою CD4 розпізнавати тільки пептидні антигени і лише тоді, коли вони знаходяться у комплексі з молекулами ГКГ ІІ класу на повер хні клітин власного організма. Сама молекула CD4 реагує з молекулами ГКГ ІІ. Молекули ГКГ ІІ знаходяться на мембрані тільки певних клітин організму - дендритних, В-лімфоцитів, макрофагів, ендотелію судин. Ці клітини мають загальну назву антигенпрезентуючих клітин. Т-хелпер здатний "побачити" своїм рецептором пептидний антиген тільки тоді , коли певна антигенпрезентуюча клітина попередньо поглинула антиген, ферментативно його переробила і зв'язала із своїми молекулами ГКГ ІІ.
Існує ще одна різновидність Т-клітин - Т-контрсупресори. Ці клітини здатні попереджувати інактивацію Т-хелперів супресорними системами. Наступною дуже важливою групою Т-лімфоцитів є цитотоксичні Т-клітини (Т-ефектори, кілери) з фенотипом CD8+TcRab. Вони здатні розпізнавати чужорідний антиген у комплексі з власними молекулами ГКГ першого класу і знищувати його. Молекула CD8 на клітинній мембрані лімфоцита взаємодіє з молекулами ГКГ І класу, які розміщені на поверхні клітин, що несуть антиген. Таким чином Т-кілери здатні також розпізнавати свій антиген за умови, що він зв'язаний з ГКГ І класу на клітинах власного організму. Молекули ГКГ І присутні на всіх клітинах організму, тому для Т-ефектора антигенпрезентуючою клітиною може стати будь-яка клітина організму, на якій розмістились білки вірусів, бактерій, найпростіших, що проникли в організм. Т-ефектори після антигенної стимуляції стають зрілими Т-кілерами, які знищують клітини - мішені, що уражені цими паразитами. Як ми бачимо, Т-лімфоцити з рецептором TcRab i CD4+ i CD8+ здатні взаємодіяти з антигеном тільки тоді, коли його їм піднесли (презентували) інші клітини. Зате вони відзначаються високою специфічністю і реагують тільки із своїм антигеном, не звертаючи уваги на подібні. Tgd-лімфоцити. Т-лімфоцити з рецептором TcRgd значною мірою відрізняються від тимоцитів з рецептором TcRab за своїми антигенрозпізнавальними властивостями. Tgd лімфоцити потрібні організму на бар'єрах (вони локалізуються у шкірі і слизових) для швидкого зв'язування антигенів без попередньої участі у цьому процесі антигенпрезентуючих клітин. Рецептор цього лімфоцита самостійно реагує з антигеном, не вимагаючи контакту з антигенами ГКГ. У той же час специфічність Tgd значно менша ніж у попередніх субпопуляцій. Один і той же лімфоцит може реагува ти з досить широким спектром антигенів. Ця фракція Т-лімфоцитів в основному реагує з інфекційними і харчовими антигенами. За попередніми даними серед Тgd можна виділити кілери, Th1,Th2.
Т-лімфоцити CD4-CD8-TcRab. Такі лімфоцити нещодавно відкриті у периферичних тканинах і відрізняються від інших тим, що здатні реагувати з ліпогліканами і сполуками міколової кислоти, які зв`язані з молекулою CD1 на поверхні антигенпрезентую чих клітин.
У лімфоїдній системі визначені спеціальні місця, в яких лімфоцити зустрічаються з антигенами. Для екзогенних антигенів, які проникають в організм через покривні бар'єрні органи, такими місцями є регіонарні лімфовузли. Для антигенів, які попадають через кров - це селезінка. Для антигенів, які проникають через бар'єр шлунково-кишкового тракту - це лімфатичні вузли брижі.
Інтенсивність рециркуляції лімфоцитів через кров'яне русло досить значна: через один лімфатичний вузол за одну добу проходить приблизно 25x109 лімфоцитів. Щоб лімфоцит попав у певний лімфоїдний орган або у певну нелімфоїдну тканину, на його клітинній мембрані присутні відповідні молекули (homing-рецептори) . Наприклад, homing-рецептор лімфоцитів, необхідний для їх попадання у лімфатичний вузол, представлений мембранною молекулою L селектина. У той же час на ендотеліальних клітинах посткапілярних венул того органа, куди повинен попасти цей лімфоцит його чекають теж конкретні молекули, комплементарні до певного homing-рецептора. Ці молекули на ендотелії судин називаються адресинами. Адресин для L селектина є молекула Gly CAM-1 (Glycan-bearing Cell Adhesion Molecule).
Гени, які кодують молекули імуноглобулінів і гени, які кодують молекули TcR, мають одну унікальну властивість якою вони відрізняються від інших клітин. Вони здатні до рекомбінації. На таких перебудованих генах синтезуються лише молекули білків TcR та імуноглобулінів. У крові людини на долю Т-лімфоцитів припадає біля 75%, 15% складають В-лімфоцити і 10 % нульові (К) клітини.
Нижче подаємо порівняльну характеристику Т- і В-лімфоцитів.
Таблиця 2
Характеристика Т- і В-лімфоцитів
ОЗНАКИ |
Т-КЛІТИНИ |
В-КЛІТИНИ |
Місце утворення |
Тимус |
Кістковий мозок (у людини) |
Густина імуноглобулінових дермі-нант |
Низька |
Значна |
Рецептори до: Fc-фрагменту IgM Гаптену Носія С3 |
Присутні у супресорів Відсутні Присутні Присутні у кілерів |
Присутні Присутні Відсутні Відсутні |
Шкірні реакції |
Сповільненого типу |
Негайного типу |
Стимуляція тимозином |
Позитивна |
Відсутня |
Утворення лімфокінів |
Значне |
Має місце |
Пригнічуючі фактори |
Тимектомія, АЛС, імуран |
Амінокапронова кислота, пуроміцин |
Основна функція |
Ефекторні клітини в ГСТ, РТПГ, трансплантаційний імунітет, хелперна і супресорна функція, продукція лімфокінів |
Попередники антитілопродуцентів |
Прояви при вродженій недостатності |
Синдром Ди-Джорджа, синдром Луї-Барра |
Агамаглобулінемія |
Дослідження кількості і функціональної активності лімфоцитів
Велике значення для дослідження стану імунної системи має підрахунок Т- і В-лімфоцитів і визначення їх функціональної здатності. Для підрахунку цих клітин з початку 70-х років вживається метод спонтанного розеткоутворення. Розеткоутворення - це процес взаємодії лімфоцитів і ксеногенних еритроцитів з утворенням клітинних конгломератів, які складаються з лімфоцита і приєднаних до нього чужорідних еритроцитів. Еритроцити розміщуються навколо лімфоцита, і на вигляд такі конгломерати нагадують розетки. Спонтанне розеткоутворення спостерігається між лімфоцитами і еритроцитами певних видів тварин.
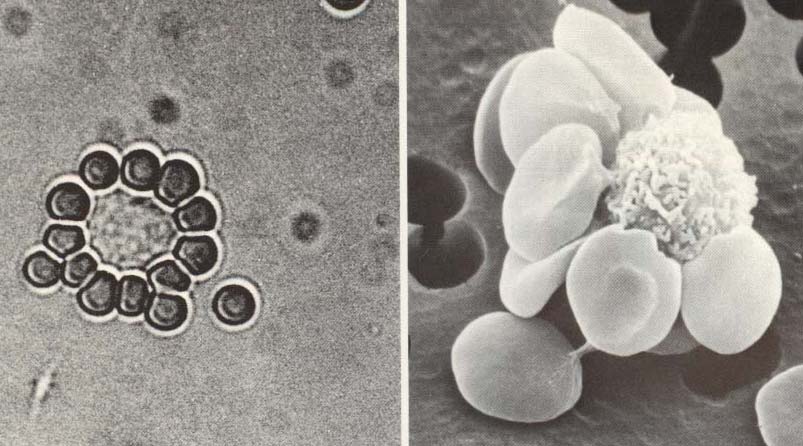
Т-лімфоцити людини, маючи рецептори до баранячих еритроцитів, утворюють з ними розетки. Ось чому метод спонтанного розеткоутворення з еритроцитами барана (метод Е-розеткоутворення - від еритроцити (Е) використовується для виявлення Т-клітин людини. Ці лімфоцити позначають як Е-РУК (Е-розеткоутворюючі клітини) (рис. 7.12).
|
Рис. 7.12. Е-розеткоутворюючі клітини (Т-лімфоцити). |
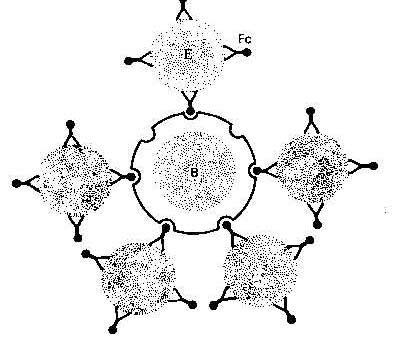
В-лімфоцити людини мають на своїй поверхні рецептори до еритроцитів мишей і формують з ними спонтанні розетки. Цей тест можна використати для підрахунку В-лімфоцитів. Проте для виявлення В-лімфоцитів більш широко до цього часу використову ються методи розеткоутворення з урахуванням інших рецепторів, які є специфічними маркерами. Такими маркерами є рецептори до Fc-фрагменту імуноглобуліну і до С3 компонента комплементу. При зв'язуванні комплекса антиген - антитіло (еритроцити - антиеритроцитарні антитіла) з рецептором до Fc фрагменту формуються так звані ЕА-розетки (рис. 7.13). Коли ж до рецептора С3 приєднується комплекс антиген - антитіло - комплемент (еритроцити - антиеритроцитарні антитіла - С3) утворюються ЕАС-розетки. Відповідно В-лімфоцити часто позначають як ЕА-РУК або ЕАС-РУК.
|
Рис. 7.13. Схема ЕА-розеткоутворення: В - В-лімфоцит; Е - еритроцит; Fc - Fc-фрагмент імуноглобуліна. |
Для вивчення функціональної здатности лімфоцитів використовується реакція бласттрансформації (РБТЛ). Перехід малих лімфоцитів із стану спокою в бластні форми, які здатні до проліферації і подальшої диференціації, називається бласттран сформацією і супроводжується морфологічними змінами - збільшенням розмірів, кількості мітохондрій, рибосом і ін. Під час трансформації в бласти в лімфоцитах стимулюються біохімічні процеси, що призводить до інтенсифікації синтезу білка, РНК, ДНК, в результаті чого відбувається мітотичний поділ клітин. Одна бластна клітина може дати клон із 32- 64 клітин, які мають ту ж імуноспецифічність, що і вихідна клітина.
Бласттрансформація лімфоцитів може бути викликана як специфічними, так і не специфічними стимуляторами. Речовини, які стимулюють мітоз лімфоцитів (мітогени) різноманітні за походженням і хімічним складом. Найбільшу їх кількість виявлено серед бактерій. Це окремі компоненти капсульної речовини, джгутиків, клітинної стінки, цитоплазматичної мембрани, рибосом. До мітогенів відносять білок А стафілококу, ліпополісахариди грамнегативних бактерій, туберкулін і т.ін.
Викликати бласттрансформацію лімфоцитів можуть окремі речовини тваринного і рослинного походження (фітогемаглюти нін - ФГА, КонА, мітоген лаконосу - РwМ).
Неспецифічні стимулятори залучають у процес бласттрансфор мації значну частину лімфоцитів незалежно від їх імунологічної специфічності. Причому одні з них вибірково активують тільки Т-клітини, інші - В - клітини. Найчастіше в клінічній практиці використовують ФГА, КонА, ЛПС, мітоген лаконосу, туберкулін. Такі мітогени, як ФГА, КонА активують тільки Т-клітини; ЛПС - тільки В-клітини.
Специфічними стимуляторами бластогенезу є антигени, які залучають у цей процес і Т-, і В- лімфоцити. На відміну від неспецифічних стимуляторів антиген здатний активувати тільки ті лімфоцити, які несуть специфічні до нього рецептори. Здатність клітин до бласттрансформації відображає функціональну активність імунокомпетентних клітин.
РБТЛ під впливом антигенів служить критерієм специфічної реактивності Т- і В- лімфоцитів, а під впливом неспецифічних стимуляторів - показником загальної імунологічної реактивності, що характеризує потенційну здатність імунної системи реагувати на антигени.