

Содержание
Исторические сведения об открытии……………………………………….2стр.
Химическая природа…………………………………………………………3стр.
Биосинтез хранения и секреции……………………………………………..7стр.
Тиреоидные гормоны………………………………………………….7стр.
Биосинтез тирогобулина……………………………………………..10стр.
Захват йодида…………………………………………………………12стр.
Органификация йодида……………………………………………....14стр.
Конденсация йодаминокислот с образованием йодтиронинов……15стр.
Секреция. Морфологический путь…………………………………..16стр.
Регуляция секреции…………………………………………………..16стр.
Ауторегуляторные эффекты йодида………………………………...18стр.
Транспорт гормона…………………………………………………………..20стр.
Биохимические функции, молекулярный механизм действия…………...22стр.
Гормональный биоритм…………………………………………………….27стр.
Болезни и лечение эндокринного расстройства…………………………..28стр.
Список литературы………………………………………………………….47стр.
Исторические сведения об открытии
В настоящее время имеется множество железистых препаратов, множество препаратов гормонов всех желез внутренней секреции, незаменимых во врачебной практике. Оказалось, что гормоны, взятые от различных животных, действуют одинаково, так же как гормоны, взятые у животного и у человека.
В то время, когда еще не знали ни наименований, ни назначения гормонов, внимание ученых привлек гормон щитовидной железы. В 1884 г. бернский хирург Теодор Кохер опубликовал отчет о произведенных им операциях на зобе. Асептика и остановка кровотечения шагнула уже столь далеко, что можно было отважиться и на такого рода операции. Кохер первый решился на это. В своем отчете он сообщал не только об успешных операциях, но и о том, что на некоторых людей они производили губительное действие: лицо опухало, физические и духовные силы убывали, наступало состояние, которое Кохер называл kachexia strumipriva, т. е. потеря сил после удаления зоба. Что происходило при этой операции? Удаляя зоб, следовательно, и щитовидную железу, он наблюдал, что при этом тело явно лишалось одного из своих важнейших органов. Но что же делает щитовидная железа в теле? Кохер и некоторые другие думали, что она служит своего рода фильтром против ядов, т. е. что это орган, который очищает тело от ядовитых веществ, и быть может, чем-то похожий на почку, но в другом роде.
То, что узнал Кохер, одновременно узнал и Мориц Шифф из Франкфурта-на-Майне — участник революции 1848 г., высланный из Геттингена и нашедший в Швейцарии не только убежище, но и исследовательскую лабораторию. Удаляя щитовидную железу у животных, он наблюдал то же, что наблюдал и Кохер у некоторых больных, — гибель живого существа от истощения всех сил.
Это противоречило тому, что физиологи говорили всего лишь несколько лет назад. «Нет даже ни одной гипотезы, касающейся функции щитовидной железы», — вот слова из одного учебника семидесятых годов. Однако результаты удаления этой железы не могли быть всегда одинаковыми потому, что она не является одиночным ясно очерченным органом, находящимся на известном месте шеи. Часто есть еще маленькие щитовидные железы, расположенные где-нибудь на другой стороне тела, действия которых оказывается достаточно для организма в том случае, если удалена основная щитовидная железа. Отсюда легко объясняется противоречие между тем, чему верили еще в семидесятых годах, и данными, собранными к 1884 г. Таким образом, если бы у больных, которым повредили операции, где-нибудь была еще одна маленькая дополнительная щитовидная железа, то и эти операции дали бы хорошие результаты.
Если удалить щитовидную железу у молодого животного, то оно отстанет в росте, его половые железы перестанут развиваться, оно застынет на той ступени физического развития, которая у человека называется идиотией; кретинизм, встречающийся иногда в горных странах, связан с недостаточностью щитовидной железы вследствие зобного перерождения этого органа. Причина такого перерождения — недостаток йода в пище или воде. У взрослого человека, у которого нет щитовидной железы или же она не функционирует, лицо опухает (это называется микседемой), отмечается появление признаков идиотии, потеря сил, ожирение.
Действующее вещество — гормон щитовидной железы, названный тироксином, открыт в 1914 г. Э. Кендаллем. Его можно изготовить и искусственно. Тироксин повышает основной обмен, способствует более интенсивному распаду белков и жиров и воздействует на углеводный обмен. Он имеет особое значение для молодых особей, так как влияет на рост костей совместно с другими гормонами с гормонами половых желез и гипофиза, передней доли железы мозгового придатка. О совместном действии гормонов, о том, что они, так сказать, образуют симфонический оркестр, в котором дирижером является гипофиз, будет рассказано далее.
Весьма часто щитовидная железа работает слишком усиленно. Впервые описанная Карлом Адольфом Базедовым в Мерзебурге базедова болезнь, симптомы которой часто приписывали всего лишь нервозности, является следствием гипертиреоза — чрезмерного повышения функции щитовидной железы.[7]
Химическая природа
В 1895 г. Магнус-Леви показал, что экстракты из щитовидной железы при введении их в организм повышают интенсивность процессов обмена веществ. Они повышают потребление кислорода у больных микседемой, у которых понижена функция щитовидной железы, а также у здоровых людей. Исследования с удалением щитовидной железы у животных и опыты с пересадкой железы оперированным животным дали прямые указания на то, что в щитовидной железе образуется активное вещество — гормон, необходимый для нормальной жизнедеятельности человека и животных. Возник вопрос о химической природе этого гормона. В конце прошлого столетия в составе щитовидной железы был открыт йод. Далее оказалось, что йод находится в железе в составе органических соединений. Полученные из ткани железы препараты тиреоглобулина содержат от 0,5 до 1 % йода. Наряду с тиреоглобулином из щитовидной железы, вернее из ее щелочных гидролизатов, было выделено йодсодержащее кристаллическое вещество — тироксин. В 1929 г. была установлена химическая структура тироксина, подтвержденная затем синтезом. Йод тироксина составляет 20—25% всего йода щитовидной железы. Веществом, близким к тироксину, в молекуле которого имеются три атома йода, является трийодтиронин. (Рис.1)
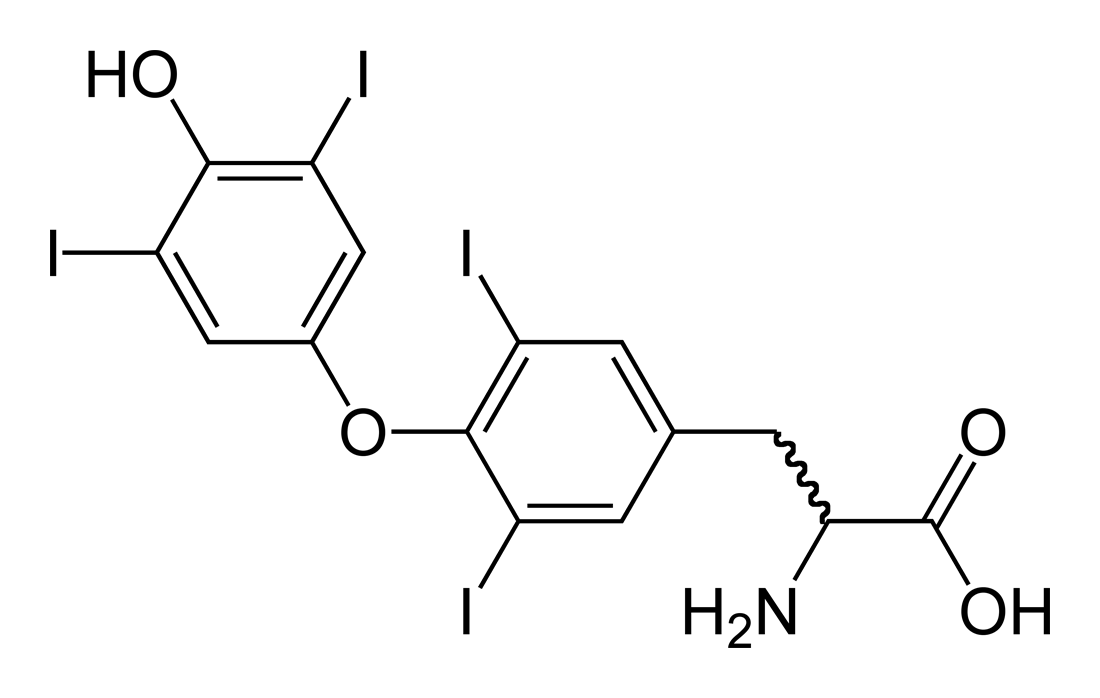
Рисунок 1. Химическая структура тироксина
Веществом, близким к тироксину, в молекуле которого имеются три атома йода, является трийодтиронин. (Рис.2)
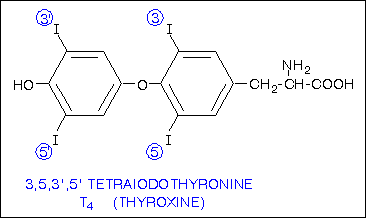
Рисунок 2. Химическая структура трийодтиронина.
Трийодтиронин выделен сравнительно недавно из гидролизатов ткани щитовидной железы и из плазмы крови.[1,том-3, C.367] Он оказался в опытах на мышах и при лечении больных значительно активнее тироксина. Установлено, что Трийодтиронин имеется в больших количествах, чем тироксин, во фракции йодсодержащих белков плазмы крови. Отсюда возникло предположение, что истинным гормоном щитовидной железы является не тироксин, а трийодтиронин. Из гидролизатов щитовидной железы выделены еще два йодсодержащих вещества: монойод- и дийодтирозин. Около 70% йода щитовиднойжелезы представлено в виде дийодтирозина. Легко заметить, что все четыре йодсодержащие вещества, полученные из гидролизатов ткани щитовидной железы, являются производным тирозина. Итак, тирозин — предшественник йодсодержащих веществ щитовидной железы. Трийодтиронин синтезируется из монойод- и из дийодтирозина или же возникает из тироксина при отщеплении от него йода. (рис.3)
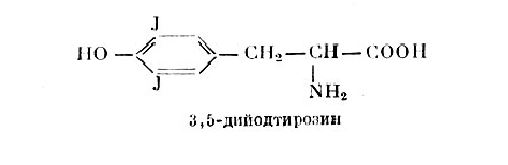
Рисунок 3. Химическая структура дийодтирозина.
В пользу предположения, что дийодтирозин является предшественником тироксина, свидетельствуют следующие факты: дийодтирозин при рН-7 в присутствии кислорода вне организма превращается в тироксин, далее он в сравнительно большом количестве имеется в щитовидной железе; физиологическое действие его невелико и он, следовательно, не может считаться гормоном щитовидной железы. Несмотря на то, что химическая природа йодсодержащих веществ щитовидной железы изучена, процессы, обеспечивающие йодирование тирозина и синтез тироксина и трийодтиронина, остаются еще не выясненными. Несомненно, одно, что щитовидная железа обладает способностью извлекать йод из притекающей к ней крови и интенсивно его включать в состав тирозина и его производных. Применение в экспериментах радиоактивного йода дало возможность выявить новые моменты, касающиеся синтеза йодсодержащих веществ в щитовидной железе. Прежде всего, была установлена связь между функциональным состоянием щитовидной железы и ее способностью поглощать из крови (концентрировать) поступающий в организм йод. При пониженной функции железы йод в ней концентрируется медленнее, наоборот, при повышенной функции йод концентрируется интенсивнее, чем обычно. Это обстоятельство используется в клинических исследованиях для диагностики функционального состояния железы. Поступивший в щитовидную железу йод быстро переходит в органическую форму. Он обнаруживается сначала в дийодтирозине, а затем в тироксине тиреоглобулина. Интересно отметить, что введенный в организм дийодтирозин не поглощается щитовидной железой и выводится из организма с мочой. Срезы ткани щитовидной железы способны синтезировать дийодтирозин и тироксин. Обязательным условием для этого является потребление ими кислорода, т. е. дыхание. Факторы, прекращающие дыхание срезов ткани железы, приостанавливают синтез дийодтирозина и тироксина. Эти данные указывают, что синтез гормона щитовидной железы связан с происходящими в ней процессами обмена веществ. [6 C.380]
Большой интерес представляет вопрос, какое из йодированных веществ, образующихся в щитовидной железе, обладает гормональной активностью. Принято считать, что этой активностью обладают тироксин и трийодтиронин, которые поступают из щитовидной железы в кровь, связываются белками сыворотки крови, а затем - доставляются к различным органам и тканям. Следует, однако, учитывать, что гормоны щитовидной железы подвергаются расщеплению с образованием промежуточных продуктов. Эти продукты могут обладать той или иной активностью. Установлено, что трий-одтируксусная кислота и трийодтирамин, возникающие из трийодтиронина, усиливают дыхание, причем действуют более эффективно, чем трийодтиронин.
Гормоны щитовидной железы обладают разносторонним действием и это, по всей вероятности, связано с тем, что из них образуются различные вещества, каждое из которых обладает определенным гормональным действием.
Установлено, что дийодтирозин тормозит образование в передней доле гипофиза тиреотропного гормона, активирующего деятельность щитовидной железы. В связи с этим дийодтирозин получил применение при заболеваниях, вызываемых гиперфункцией щитовидной железы.
Биосинтез хранения и секреции
Тиреоидные гормоны.
Клеточные и молекулярные механизмы, используемые щитовидной железой для биосинтеза, накопления и секреции тиреоидных гормонов в течение прошлых 3—4 десятилетий, изучены достаточно полно благодаря объединенным усилиям большого числа исследователей. Подробности этих исследований содержатся в ряде исчерпывающих обзоров. Синтез тиреоидных гормонов во многих. отношениях сходен с синтезом полипептидных и белковых гормонов с добавлением нескольких уникальных этапов, особенно подходящих к метаболическим функциям щитовидной железы.[7]
Основными тиреоидными гормонами являются йодтиронины — тетрайод-L-тиронин (тироксин, Т4) и 3,5,3-трийод-L-тиронин (трийодтиронин, Т3), которые образуются из йодированных тирозиновых предшественников. Они состоят из йодтирозина (моно — или дийодированного) и йодированного фенольного кольца, объединенных 4,1-эфиpнoй связью. Эти два йодтиронина вместе с йодтирозиновыми предшественниками составляют примерно 90% органического или белковосвязанного йода в щитовидной железе, причем на долю Т4 и Т3 приходится 99% секретируемого органического йода. Процессы образования и секреции тиреоидных гормонов включают несколько этапов, часть которых отличается своеобразием и характерна только для щитовидной железы. .[7]
Главными веществами, используемыми в синтезе тиреоидных гормонов, являются йод и тирозин. Щитовидная железа отличается высокоэффективным механизмом захвата йода из крови, а в качестве источника тирозина она синтезирует и использует крупный гликопротеин — тироглобулин. Если тирозин в организме содержится в большом количестве и поступает как из пищевых продуктов, так и из распадающихся эндогенных белков, то йод присутствует лишь в ограниченном количестве и поступает только из пищевых продуктов. Йод всасывается из кишечника в форме йодида и в этой форме циркулирует в крови в свободном (несвязанном) состоянии. .[7]
Йодид, захватываемый из крови тиреоидными (фолликулярными) клетками, и тироглобулин, синтезируемый в этих клетках, секретируются (путем экзоцитоза) во внеклеточное пространство внутри железы, называемое просветом фолликула или коллоидным пространством, окруженное фолликулярными клетками. В просвете фолликула или (что более вероятно) на апикальной поверхности клеток, обращенной в этот просвет, йодид окисляется в йод и другие окисленные продукты и ковалентно связывается фенольными кольцами тирозиновых остатков, содержащихся в полипептидном каркасе тироглобулина. Связывание йода с фенольным кольцом происходит либо только в 3-м положении, либо как в 3-м, так и в 5-м положениях, в результате чего образуются монойодтирозин (МИТ) и дийодтирозин (ДИТ) соответственно. Этот процесс йодирования тирозиновых остатков тироглобулина известен под названием этапа органификации в биосинтезе тиреоидных гормонов. .[7]
Следующим этапом является конденсация йодтирозинов с образованием йодтиронинов. Все еще оставаясь в структуре тироглобулина, молекулы МИТ и ДИТ конденсируются, образуя трийодтиронин (Т3), и подобно этому две молекулы ДИТ конденсируются, образуя молекулу L-тироксина (Т4). В таком виде, т. е. связанные с тироглобулином, йодтиронины, равно как и непроконденсировавшие йодтирозины, хранятся в тиреоидном фолликуле. Этот комплекс йодированного тироглобулина часто называют коллоидом. Таким образом, тироглобулин, составляющий 10% от влажной массы щитовидной железы, служит белком-носителем, или предшественником накапливающихся гормонов. .[7]
Секреция тиреоидных гормонов — процесс, происходящий в ответ на метаболические потребности и опосредуемый действием тиротропного гормона (ТТГ) на тиреоидные клетки, — предполагает высвобождение гормонов из тироглобулина. Это происходит на апикальной мембране путем поглощения коллоида, содержащего тироглобулин (процесс, известный под названием эндоцитоза). Тироглобулин затем гидролизуется в клетке под влиянием протеаз, а высвобождаемые таким образом тиреоидные гормоны выделяются в циркулирующую кровь. (Рис.4)
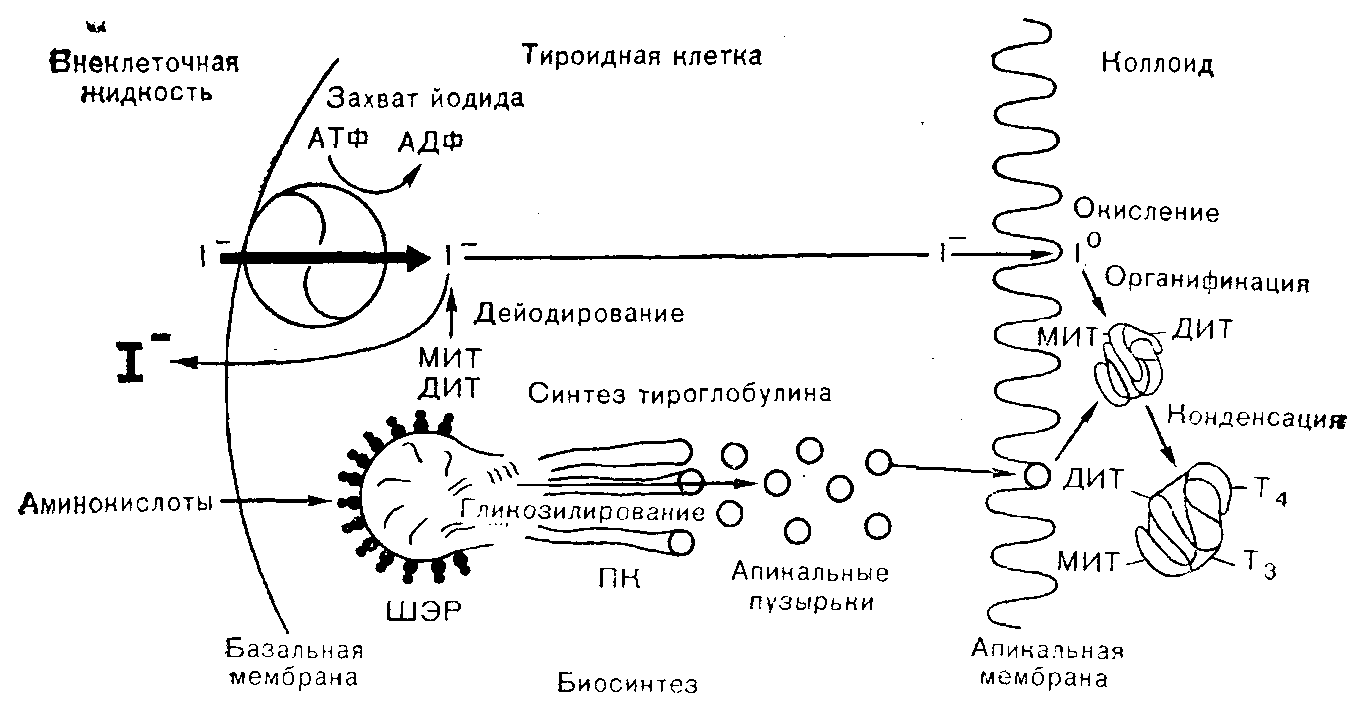
Рисунок 4. Схематическое изображение пути биосинтеза тиреоидных гормонов в клетке. Т3 и Т4 образуются и хранятся связанными с тироглобулином в коллоиде.
Биосинтез
Различные этапы биосинтеза и секреции тиреоидных гормонов L-трийодтиронина (Т3) и L-тироксина (Т4) следует рассмотреть отдельно и более подробно, подразделив их на следующие категории: 1 – биосинтеза тироглобулина; 2 — захват йодида, 3 — органификация йодида; 4—конденсация; 5—поглощение клетками и протеолиз коллоида; 6 — секреция.
Биосинтез тироглобулина.
Тироглобулин представляет собой крупный гликопротеин с молекулярной массой примерно 670 000 (19S). Он состоит по меньшей мере из 4 субъединиц (из двух сходных пар разных субъединиц), связанных друг с другом комбинацией нековалентных и ковалентных. (дисульфидных) связей. Белковый комплекс содержит от 8 до 10% углеводов. Тироглобулин состоит примерно из 5900 аминокислотных остатков, около, НО из которых—тирозиновые. Содержание йода в тироглобулине, полученном из интактных желез, колеблется от 0,2 до 1% и более. Тироглобулин синтезируется на полирибосомах, связанных с ШЭР. Субклеточный путь, по которому транспортируется вновь синтезированный тироглобулин, характерен для транспорта многих предназначенных к секреции белков. Вновь синтезированные полипептидные цепи направленно проникают через мембрану ретикулума во внутрицистернальное пространство ШЭР, по которому переносятся в пластинчатый комплекс и упаковываются в окруженные мембраной пузырьки, называемые апикальными пузырьками (См. рис. 3и 4). Эти пузырьки транспортируются к апикальному краю клетки, где сливаются с мембраной, граничащей с фолликулярным, или коллоидным, пространством, и тироглобулин выбрасывается в это пространство. Именно на поверхности этой мембраны со стороны коллоида и йодируется тироглобулин. Исследования с импульсной меткой и последующим ее разведением, проводимые на срезах щитовидной железы, инкубируемых с радиоактивными аминокислотами in vitro, показали, что процессы синтеза и транспорта тироглобулина (от момента начального синтеза до первого появления в коллоидном пространстве) занимают от 4 до 6 ч. Начальным продуктом синтеза является, по-видимому, субъединица с молекулярной массой около 160000 (3—8S). Предполагается, что разные субъединицы кодируются отдельными мРНК. (Рис.5)
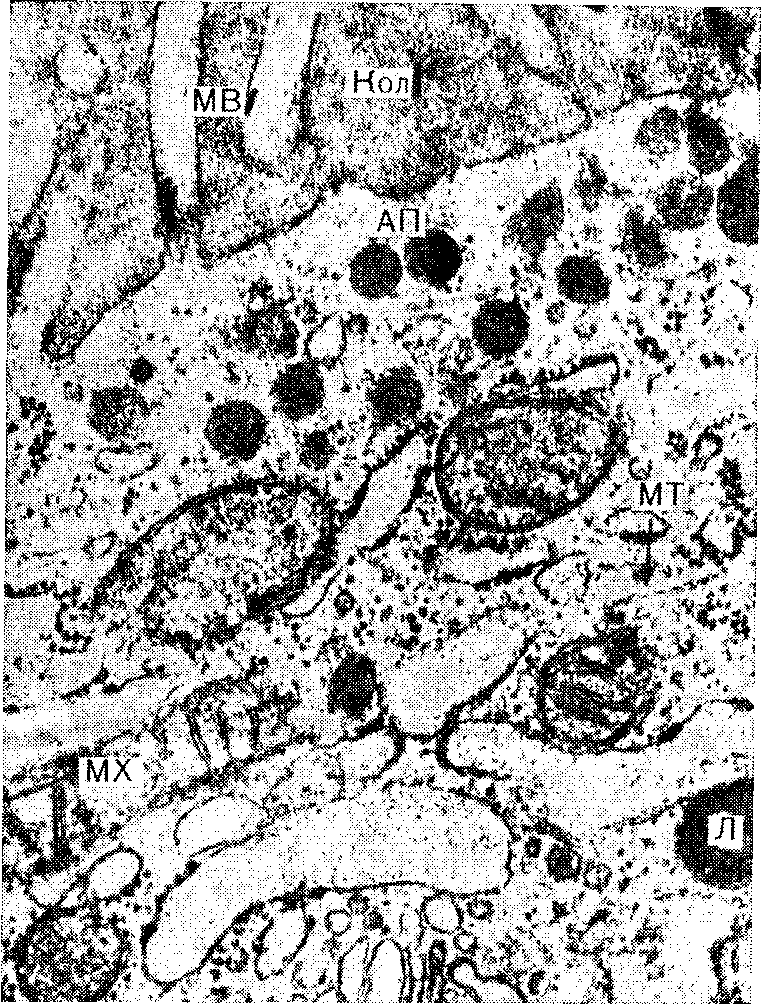
Рисунок 5. Участок тиреоидной фолликулярной клетки. Вверху можно видеть микроворсинки (MB), тянущиеся от апикального края клетки в коллоид (Кол), в цитоплазме под краем — апикальные пузырьки (АП).
Плотное тельце, имеющее лизосомную (Л) природу; МТ — микротрубочки, MX — митохондрия Электронная микрофотография. х37 800.
Затем субъединицы объединяются в димеры с молекулярной массой 320 000 (12S,) и образуют нейодированный тироглобулин (17S), который «созревает» во время и после йодирования в окончательный комплекс с молекулярной массой 667000 (19S). Последовательность такова: 4 субъединицы (по 3—8S) 2 субъединицы (по 12S) нейодированный («незрелый») тироглобулин (17S) йодированный («зрелый») тироглобулин (19S). Гликозилирование тироглобулина происходит на всех стадиях синтеза. Сердцевинный олигосахаридный комплекс присоединяется, вероятно, к отдельным аспарагиновым остаткам N-гликозидной связью и переносится с долихофосфатного носителя в ходе транспорта вновь образуемых цепей субъединиц через мембраны эндоплазматической сети. В процессе транспорта полипептида по цистернам эндоплазматического ретикулума в пластинчатый комплекс к олигосахаридным ядрам добавляются новые углеводные остатки. По всей вероятности, гликозилирование каким-то образом участвует в транспорте и сборке цепей субъединиц и необходимо для формирования полностью собранного тироглобулинового комплекса с его специфической трехмерной четвертичной структурой. .[7]
Захват йода.
Важнейшим этапом биосинтеза тиреоидных гормонов является активный, контролируемый ферментами, транспорт йодида, содержащегося в крови, через мембрану тиреоидной клетки в цитозоль и из него в фолликулярный коллоид. В конечном счете, обеспеченность организма йодом должна определяться пищевыми продуктами. Часть йода всасывается в органической форме и превращается в йодид в печени. Однако наибольшая его часть уже. всасывается в форме йодида и именно в этой форме разносится с кровью. Суточное потребление йодида с пищей в США составляет примерно 500 мкг. Из этого количества 120 мкг поглощается щитовидной железой и 60 мкг включается в тиреоидные гормоны и секретируется с Т4 и Т3. Поглощение и концентрирование йодида щитовидной железой обеспечивается двумя энергозависимыми контролируемыми ферментными «насосами», или механизмами захвата; один, расположенный на базальной, или капиллярной, мембране, извлекает йодид из крови и транспортирует его в цитозоль тиреоидной клетки, а второй, локализующийся на апикальной мембране, доставляет его в коллоидное пространство. Существует и конкурентный пассивный перенос йодида обратно в кровь, но «чистый» эффект сводится к концентрированию йодида в щитовидной железе. Эти насосы функционируют столь эффективно, что концентрация йодида в щитовидной железе не менее чем в 25 раз превышает его уровень в плазме крови, а в условиях дефицита йода концентрация йодида в железе может быть в 500 paз выше, чем в плазме. Эффективность, с которой щитовидная железа концентрирует йодид, находится в обратной зависимости от уровня доступного йодида в плазме, и в условиях относительной йодной недостаточности железа извлекает до 20% йодида, присутствующего в крови. Извлеченный тиреоидной клеткой из крови йодид быстро уравновешивается в общем пространстве йодида, уже содержащегося в клетке, который образуется в результате распада и дейодирования йодаминокислот, высвобождающихся из-тироглобулина (см. далее), и быстро проникает в коллоидное-пространство, где связывается с тироглобулином. Этот концентрирующий и транспортный механизм работает так быстро, что отношение «свободного» йодида к органически связанному в щитовидной железе может составлять лишь 10:8000 мкг. Авторадиографические исследования показали, что уже через 30 с после импульсного внутривенного введения животному радиоактивного йодида практически вся метка обнаруживается в коллоидном пространстве, будучи органически связанной с тироглобулином ; только около 0,25% тиреоидного йода остается в форме свободного йодида, а остальная его часть связана с тироглобулином в форме йодаминокислот или йодтиронинов. .[7]
О необходимости столь легкого накопления йода в щитовидной железе свидетельствует тот факт, что для поддержания нормальной секреции гормонов эта железа должна каждые сутки накапливать от 50 до 75 мкг йодида, т. е. полностью экстрагировать йодид из 20—30% крови, протекающей через щитовидную железу в сутки, что составляет лишь небольшую часть резерва, необходимого на случай колебаний содержания йода в диете или возникновения чрезмерной потребности в тиреоидных гормонах. Сам организм, однако, способен в значительной мере компенсировать временный дефицит йодида путем повышения его канальцевой реабсорбции и всасывания в кишечнике. .[7]
Детали конструкции йодидных насосов выяснены не полностью» но, по всей вероятности, в них участвует Na+/K+-стимулируемая, Mg2+-зависимая оуабаинчувствительная АТФаза. Для накопления йодида необходимы и ионы кальция. Процесс концентрирования явно зависит от доступности клеточной энергии, поскольку разобщители окислительного фосфорилирования, такие, как 2,4-динитрофенол и бис-оксикумарин, являются эффективными ингибиторами накопления йода срезами щитовидной железы. .[7]
Щитовидная железа концентрирует не только йодид, но и некоторые другие анионы, которые могут действовать как конкурентные ингибиторы транспорта йодида. К таким ингибиторам относятся тиоцианат, перхлорат и пертехнетат. Перхлорат можно применять в клинике для того, чтобы вызвать быстрый выброс неорганического йодида, после определения количества которого можно получить представление о величине несвязанной фракции йодида в щитовидной железе. У больных с нарушенной органификацией йодида, например при врожденном дефиците ферментов или воздействии зобогенных факторов, блокирующих органификацию йодида, одна и та же доза перхлората будет высвобождать гораздо большую, чем в норме, часть дозы ранее введенного» йодида.
Органификация йодида
На апикальной мембране тероидной фолликулярной клетки илд вблизи нее йодид окисляется пероксидазными ферментами, что приводит к присоединению йода к фенольному кольцу тирозиновых остатков, содержащихся в структуре тироглобулина. Йодирование молекул тирозина происходит именно тогда, когда они являются частью полипептидного каркаса тироглобулина, а не до синтеза этого белка. Хотя некоторые авторы обнаруживали пероксидазную активность в митохондриях и пластинчатом комплексе, современные данные свидетельствуют в пользу той точки зрения, — что основное количество пероксидазы связано с микроворсинками апикальной мембраны и что процесс йодирования происходит на поверхности раздела мембрана — коллоид, а не внутри самой фолликулярной клетки. Территориальное ограничение йодирования поверхностью раздела между клеткой и коллоидом обеспечивает щитовидную железу механизмом минимизации случайного йодирования внутриклеточных белков, не являющихся тироглобулином. В результате йодирования тироглобулин «созревает» и его константа седиментации увеличивается с 17 до 19S. Относительное количество моно- и дийодтирозинов варьирует в зависимости от степени йодирования, которая в свою очередь зависит от доступности йода. В тироглобулине человека в норме йод распределен следующим образом: 20% в составе МИТ, 40% —ДИТ, 30% —Т4 и 2% —Т3. Эти величины свидетельствуют о том, что в большинстве случаев фенольные кольца содержат две молекулы йода. Можно отметить, что в исследованиях по химическому йодированию модельного вещества — N-ацетилтирозина in vitro было установлено в 30 раз более интенсивное монойодирование, чем дийодирование; поэтому, если бы тирозиновые группы тироглобулина вели себя подобно модельному соединению, то основная часть йода в тироглобулине должна была бы присутствовать в монойодтирозинах. Однако в тироглобулине, выделенном из щитовидных желез больных зобом, доля монойодированных веществ заметно превышает таковую дийодированных (см. далее). Содержание тирозина в тироглобулине отнюдь не уникально и сравнимо с его содержанием во многих других белках (примерно 1,8% всех аминокислот). Имеются данные, указывающие на то, что подвергаться йодированию могут лишь особые тирозиновые остатки в аминокислотной последовательности тироглобулина; предполагается, что тироксин (Т4) локализуется в уникальной последовательности: ала-сер-Т4-глю (или глн)-асп (или асн) . По всей вероятности, в определении доступности отдельных тирозиновых остатков для йодирования важную роль играет вторичная и/или четвертичная структура молекулы тироглобулина. Вещества, относящиеся к классу тионамидов (метимазол, пропилтиоурацил) являются мощными ингибиторами этапа органификации йодида вследствие их тормозящего действия на пероксидазу щитовидной железы. .[7]