

3.2.4.2. Мітохондрії
У клітинах еукаріот рибосоми містяться не лише в гіалоплазмі, але і в мітохондріях (а в рослинних клітинах - ще і в хлоропластах).
Окрім рибосом, мітохондрії (і хлоропласти) містять так само свою ДНК, здатну до реплікації і транскрипції. Тобто є автономний геном і автономна система його експресії.
Ця система багато в чому схожа з такою у прокаріот:
а) ДНК є не лінійною, як в хромосомах ядра, а кільцевою;
б) рибосоми по своїх параметрах близькі до бактеріальних (см. рис. 3.8), тобто дещо дрібніші за рибосоми цитоплазми;
в) для ініціації трансляції використовується формілМет - тPHKjмет.
При цьому мітохондріальна ДНК (мтДНК) по розміру невелика: містить 15 000 нуклеотидних пар, що на 3-4 порядки менше, ніж в ДНК хромосом.
В одній мітохондрії є 1-2 молекули мтДНК; вцілому у всіх мітохондріях клітини досить багато таких молекул – від 50 до 3000. Вірогідно, що всі вони практично однакові.
Сумарна частка цих мтДНК в загальній масі ДНК клітини є досить невеликою – не більше 0,5-1,0%.
Ще менший внесок мтДНК в загальну кількість генів. Так, мтДНК кодує лише 10-20 мітохондріальних білків, що складає лише 5% всіх білків мітохондрій. Крім того, мтДНК кодує рРНК мітохондріальних рибосом і мітохондріальні тРНК.
У внутрішньохондріальній трансляції використовується всього 22 види тРНК, тоді як в цитоплазмі їх – біля 60.
Отже, вся ця автономна система забезпечує утворення досить невеликої кількості мітохондріальних білків. Останні 95% білків мітохондрій (не говорячи про всі інші клітинні і позаклітинні білки) кодуються хромосомами ядер і синтезуються цитоплазматичними рибосомами.
Що створює проблему переносу ново синтезованих білків із гіалоплазми в мітохондрії.
Виникає питання: навіщо взагалі необхідно створювати автономний мітохондріальний геном в автономну систему його експресії?
Найбільш популярна відповідь полягає в тому, що мітохондрії (і хлоропласти) – це потомки давніх прокаріот, які проникли в еукаріотичні клітини і утворили симбіоз.
3.3. Інгібітори трансляції
3.3.1. Інгібірування трансляції у бактерій
Як ми вже сказали (п. 3.2.4.1), більшість антибіотиків (речовини, виділені з грибів, бактерій і т. д.) є специфічними інгібіторами трансляції у мікроорганізмів - без помітної дії (у терапевтичних дозах) на аналогіичний процес в клітинах хазяїна. Це як би хімічне оточення, використовуване одними мікроорганізмами проти інших.
Тут ми назвемо лише деякі з величезного числа таких з'єднань. Безпосереднім об'єктом їх впливу є ті або інші функціональні центри рибосом (мал. 3.11) - у складі малої або великої субодиниці.
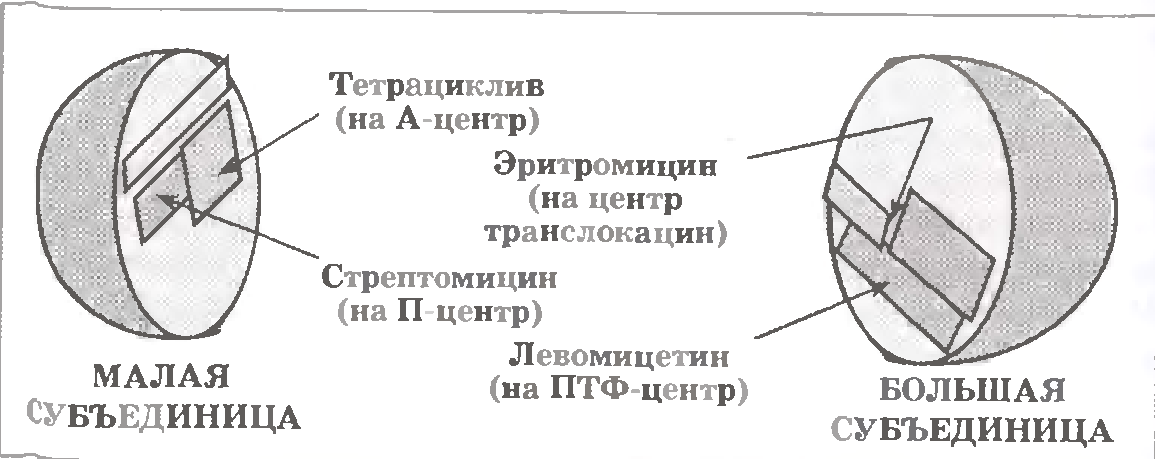
а) Антибіотики, діючі в області малої (30S ) субодиниці.
Стрептоміцин впливає на ту частину П-центру, яка знаходиться на малій субодиниці. Тим самим він утруднює зв’язування ініціаторної аа-тРНК (формілМет - тPHKjмет), тобто інгібує ініціацію білкового синтезу.
Якщо проте ініціація сталася, то все одно у присутності стрептоміцину пептидил-тРНК погано зв'язана з малою субчасткою рибосоми. Це робить рибосомний комуплекс неміцним, чому він нерідко розпадається передчасно.
Інший відомий антибіотик - тетрациклін - діє на А-центр 30S-субчастинки. Тому інгібується зв’язування чергової аа-тРНК.
б) Антибіотики, діючі в області великої (50 S ~) субодиниці.
Левоміцетин (чи хлорамфенікол) інгібірує активність ПТФ-центру.
А еритроміцин діє на ту ділянку великою субъединицы, який відповідає за транс локацію. В результаті, нова пептидил-тРНК (що утворилася після чергової ПТФазної реакції) залишається в А-центрі (не переміщуючись в П-центр) і перешкоджає зв'язуванню чергової аа-тРНК.
Усі ці ефекти, як вже двічі відзначалося, мають вибірковий характер: відносно еукаріотичних рибосом вони відсутні. Але це не означає повної нешкідливості антибіотиків; на організм пацієнта вони можуть робити немало побічних дій, хоча і іншими способами.