

Організація генетичного матеріалу у еукаріот Гени ряду білків і рнк
Тепер звернемося до еукариот.
Як ми знаємо, одна з відмінних рис більшості генів еукаріот - наявність в їх складі некодуючих ділянок - інтронів.
Інша особливість полягає в тому, що разом з унікальними генами (представленими в гаплоїдному геномі одиничним числом копій) зустрічаються гени, що багаторазово повторюються.
Щоб проілюструвати ці дві особливості, розглянемо деякі конкретні гени.
Гени гістонів
Гістони - основні (по кислотно-лужних властивостям) білки, що беруть участь у формуванні нуклеосомної структури хроматину. Кожен з п'яти видів цих білків (Н1, Н2А, Н2В, Н3 і Н4) кодується відповідним геном.
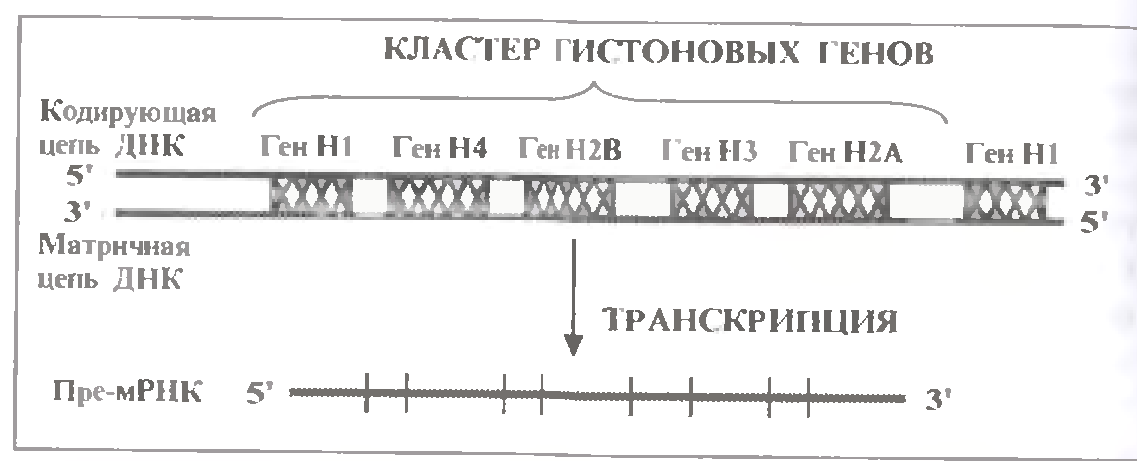
Ось характерні риси організації цих генів (див.мал):
а) всі 5 гістонових генів згруповані в єдиний кластер довжиною приблизно в 6900 н.п.
б) такі кластери повторюються в геномі багатократно: у людини – приблизно 35 раз (в гаплоїдному наборі хромосом), у морського їжака – 300-1000 раз. Це пришвидшує синтез гістонів в S- фазі клітинного циклу.
В хромосомі кластери слідують тамдемно один за одним.
в) В різних кластерах однотипні гістонові гени не завжди повністю ідентичні. Тому і самі гістони (Н1, Н2А, Н2В) дуже схожі, але в різних білках.
г) В усіх кластерах гени розташовуються в одній і тій же послідовності і розділені між собою спейсерами.
На спейсеры припадає близько 70 % усієї довжини кластера.
д) Важлива особливість гістонових генів - відсутність в них інтронів (чим вони відрізняються від більшості генів еукаріот).
Інша особливість - відносний високий вміст пар ГЦ : так, в гені Н3 воно дорівнює 0,52, а в середньому по усій ДНК людини - 0,39.
е) У людини і більшості інших еукаріот в усіх п'яти гістонових генах в якості кодуючої виступає один і той же ланцюг ДНК.
У дрозофіли, як вже відзначалося, в кластері гістонових генів в трьох випадках (гени Н1, Н2А і Н3) кодуючим є один ланцюг, а в двох інших випадках (гени Н2В і Н4) - інший ланцюг. При цьому гени розташовані в такому порядку (Н1, Н2В, Н2А, Н4, Н3), що гени з протилежною орієнтацією правильно чергуються один з одним.
ж) Виключаючи дрозофілу, у еукаріот кластер гістонових генів транскрибується як єдине ціле - у вигляді однієї довгої про-мРНК, яка містить інформацію про усі п'ять гістонові білки.
Але при дозріванні цієї про-мРНК вона розрізається на п'ять окремих гістонових мРНК.
У дрозофіли гени гістонів повинні транскрибуватися окремо, причому у різних напрямах.
Гени рибосомних РНК
Як уже згадувалося, до складу рибосом входять рРНК чотирьох видів.

Дані РНК розрізняються по константі седиментації; відповідно до цього їх і позначають:
а) Гени усіх рРНК локалізуються в ядерцевих організаторах, тобто тих ділянках хромосом, які асоційовані з ядерцями.
При цьому ген найменшої рРНК (5S-рРНК) розташовується окремо від генів інших рРНК. А гени інших трьох рРНК об'єднані в кластер (див.мал.).
б) Подібно гістоновим генам, гени рРНК представлені великим числом копій. У людини кількість копій (у розрахунку на гаплоїдний геном) приблизно дорівнює 100, у жаби в соматичних клітинах - близько 900.
У дозріваючих ооцитах жаби відбувається вибіркове і різке збільшення числа генів рРНК (т. з. ампліфікація) - до 2 млн. копій на ядро.
в) Інша схожість з гістоновими генами полягає у відсутності інтронів, а також у вищому (чим в середньому по ДНК) відносному вмісті пар ГЦ.
г) Довжина кластера трьох генів рРНК - приблизно 8000 н. п. У кластері гени розділені двома спейсерами, але особливо великим є спейсер між сусідніми кластерами - близько 5000 н. п.
д) Кожен кластер знову (подібно до кластера гістонових генів) транскрибується як єдине ціле: утворюється одна молекула про-рРНК, що містить (окрім спейсеров) послідовність усіх трьох рРНК.
Таким чином, організація генів рРНК і гістонів є дуже схожою.
Гени гемоглобіну
У нормі, як відомо, зустрічаються 4 види гемоглобіну (Нb), до складу яких входять субодиниці 5 видів:
Вид Нb |
Нb А Гемоглобін дорослих |
Нb А2 Гемоглобін дорослих |
Нb ембріона |
Нb F Нb плоду |
Субодиничний склад |
α2 β2 |
α2 δ2 |
α2 ε2 |
α2 γ2 |
Причому в Нb F-ланцюга бувають двох видів: γG і γА (відмінність - по одному амінокислотному залишку).
Відповідно, білкова частина Нb кодується шістьма генами: α, β, δ, ε, γG і γА. (Крім того, декілька генів кодують ферменти синтезу тема - небілкового компонента Нb).
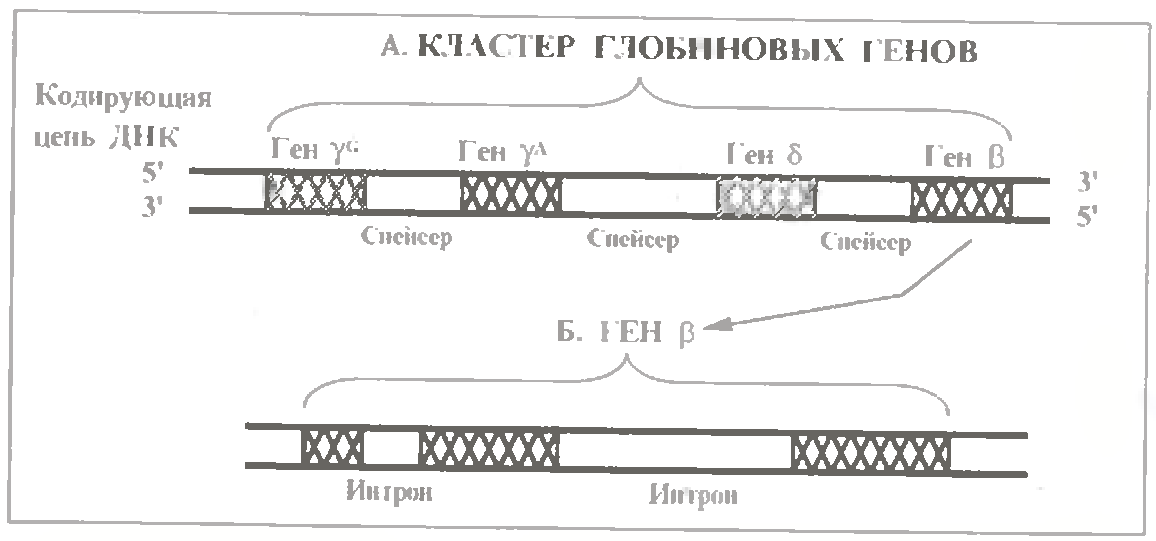
а) На відміну від попередніх генів, гени Нb вважаються унікальними: представлені дуже невеликим числом копій.
б) При цьому α-ген глобіну знаходиться в іншій хромосомі, ніж інші глобінові гени, і повторюється в цій хромосомі двічі.
А чотири глобінові гени - γG, γА, α і β - об'єднані в кластер (див.мал.), який теж повторюється лише декілька раз.
Таке розташування генів може мати два не виключаючих один одного пояснення.
По-перше, гени, що входять в кластер, мабуть, утворились в еволюції з одного гена, який кілька разів подвоювався. Після цього його копії еволюціонували кожна по-своєму.
По-друге, субодиниці, що кодуються цими генами, на відміну від α-ланцюгів, входять до складу лише якогось одного виду Нb, який утворюється на певній стадії онтогенезу. Сусідство відповідних генів може полегшувати перемикання активності з одного гена на інший.
в) В глобіновому кластері дуже великі три спейсерні ділянки: варіюючи від 4000 до 14 000 н. п., вони складають більшу частина довжини кластера.
Усередині генів містяться досить великі діялнки интронів: в гені β їх два - з довжиною 120 і 555 н. п.
г) Транскрибуються гени глобінового кластера, очевидно, окремо один від одного (оскільки їх експресія проходить на різних стадіях онтогенезу).
Чинники транскрипцій і репресори.
На функціонування генів чинять вплив більшість білків. Причому цей вплив не укладається в ті відносно прості схеми оперонної регуляції, які реалізуються у бактерій.
Так, останніми роками виявлено і описано велика кількість білків, що мають властивості факторів транскрипцій або репресорів (або тих і інших відразу). При цьому вони вступають в різноманітні взаємини один з одним, а також з іншими речовинами, від чого залежить кінцевий вплив на активність генів.
Іншими словами, подобно складної сітки із сотен метаболічних перетворень, в клітинах еукаріот є не менш складна сітка регуляторних взаємовідношень. Повної картини цієї сітки поки немає, але окремі її фрагменти вже відомі.
В данную сітку входять не тільки білки, які безпосередньо взаимодіють з ДНК, але й велика кількість інших речовин — позаклітинні сигнальні речовини (в т. ч. гормони), клітинні і внутрішньоклітинні рецептори, внутрішньоклітинні посредники (медіатори). До того ж далеко не кожна регуляторна дія має своїм кінцевим ефектом зміну активності генів.
З усіх цих причин регуляторні взаємини у еукариот вимагають окремого розгляду, який ми розглянемо пізніше.
Тут же ми коротко відмітимо тільки ті моменти, які торкаються ДНК зв'язуючих білків, що виступають чинниками транскрипцій або репресорів. Таким чином, ви повинні мати на увазі, що йтиметься лише про кінцеву ланку складних регуляторних ланцюгів.
Поділ ДНК зв'язуючих білків по їх структурі
У різних білках ДНК-зв'язуючих білків часом зустрічаються схожі структурні елементи. За цією ознакою виділяють декілька родин таких білків. Проте потрібно відмітити, що не кожен ДНК-зв'язуючий білок вдається віднести до тої або іншої родини.
а) Білки, що містять мотив "спіраль-поворот-спіраль".
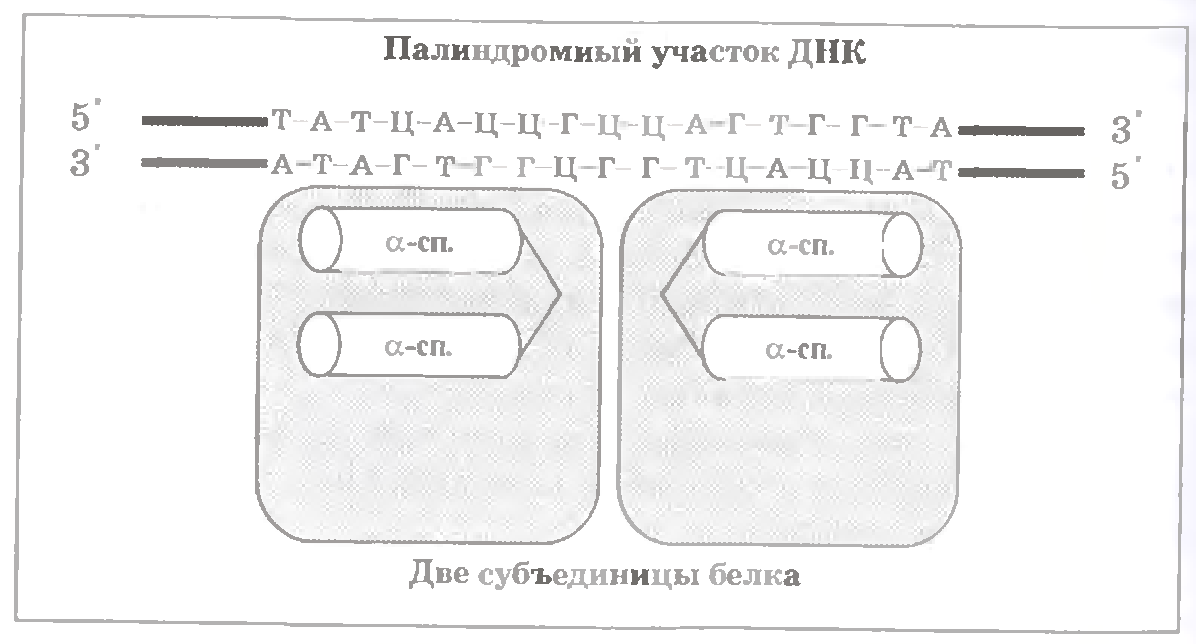
У цих білках (див.мал.) ДНК-впізнаваюча ділянка (складає невелику частину білку) включає дві α-спіралі, сполучених петлею. З них одна спіраль безпосередньо зв’язується з ДНК, специфічно взаємодіючи з визначеною послідовністю нуклеотидних пар.
У білці - дві однакові субодиниці, які одновременно беруть участь у взаємодії з ДНК. Це є можливим завдяки тому, що відповідна нуклеотидна послідовність ДНК є паліндромною, тобто, з урахуванням полярності ланцюгів, читається однаково з обох боків. (З паліндромами ми вже стикалися)
Особливо поширені подібні білки у прокаріот: маються на увазі білки-репресори.
Нижченаведені ж родини білків характерні, мабуть, лише для еукаріот.
б) Білки, що містять гомеодомени.
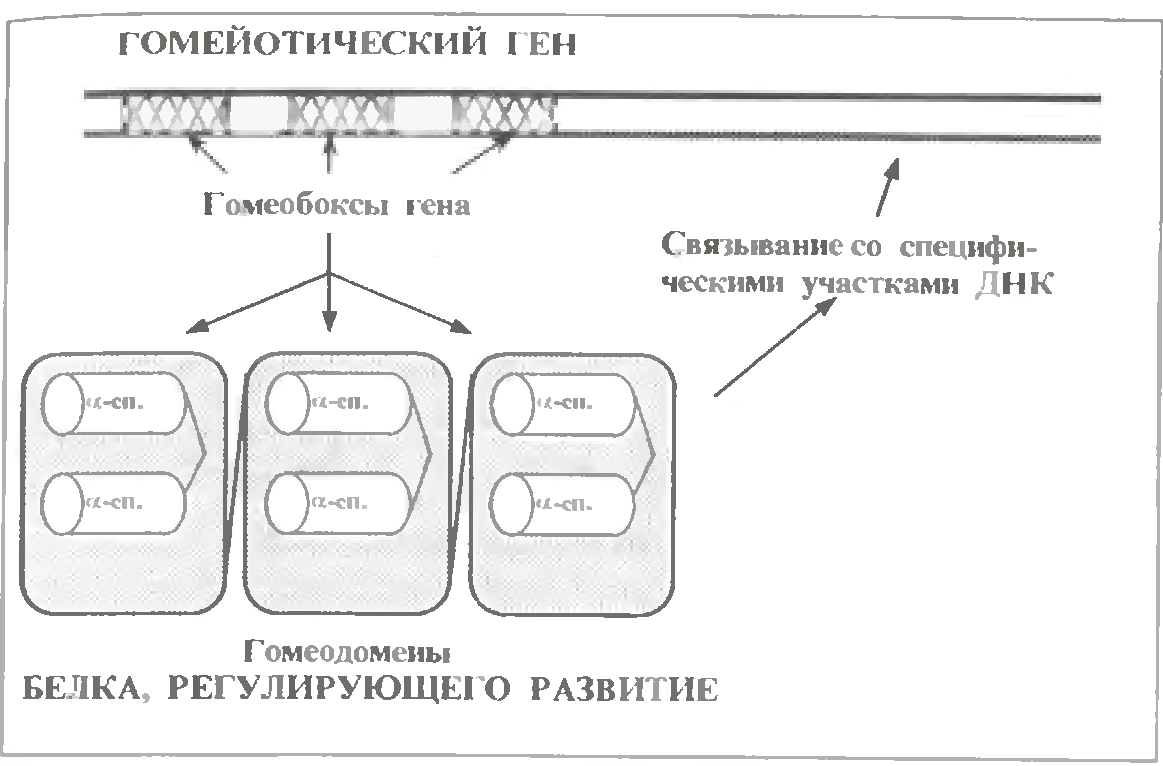
Ці білки продукти т. з. гомейотичних генів еукаріот, тобто генів, відповідальних за ембріональний розвиток.
За допомогою цих білків здійснюється керування розвитку шляхом включення одних і виключення інших генів.
Відмінна риса цих білків - наявність в них однотипних доменів, які називаються (по функції кодуючих генів) гомеодоменами (див.мал.).
Як відомо, домени - це фрагменти білкової субодиниці, що мають відносно самостійну третинну структуру. Іншими словами, в цьому випадку єдиний поліпептидний ланцюг при згортанні в третинну структуру утворює не одну велику, а дещо дрібніших глобулоподоб-них ділянок, пов'язаних між собою "лінкерними" ділянкими ланцюга.
Доменна організація зустрічається тільки у досить великих субодиниць (білків) - містять не менше 200 амінокислотних залишків. Фрагменти гена, що кодують різні домени білку, обов'язково розділені інтронами, тобто представляють собою різні екзони.
У разі гомейотических генів такі екзони називаються гомеобоксами. Отже, гомеобокс - це екзон гомейотичного гена, кодуючий гомеодомен відповідного білку.
Гомеодомен утворений приблизно 60 амінокислотними залишками. По структурі ж він нагадує белки-репресори прокаріот, тобто теж має мотив "спіраль-поворот-спіраль". І так само одна з α-спиралей цього мотиву специфічно взаємодіє з певними ділянками ДНК.
Гомеодомени різних білків цієї родини дуже схожі один з одним, що свідчить про спільність їх походження. Можливо, вони дійсно пішли від білків-репресорів прокаріот. Але при цьому кожен з гомеобілків робить дію не на окремий оперон, а на набагато більшу сукупність генів.
Дійсно, в процесі розвитку відбуваються масштабні зміни просторової структури великих фрагментів хромосом. Якісь з цих фрагментів переходять в стан гетерохроматину, інші - навпаки, в стан еухроматина.
Між іншим, звідси витікає, що і в гетерохроматині мають бути послідовності ДНК, доступні для впізнавання гомеодоменними білками.
в) Білки, що містять "цинкові пальці".
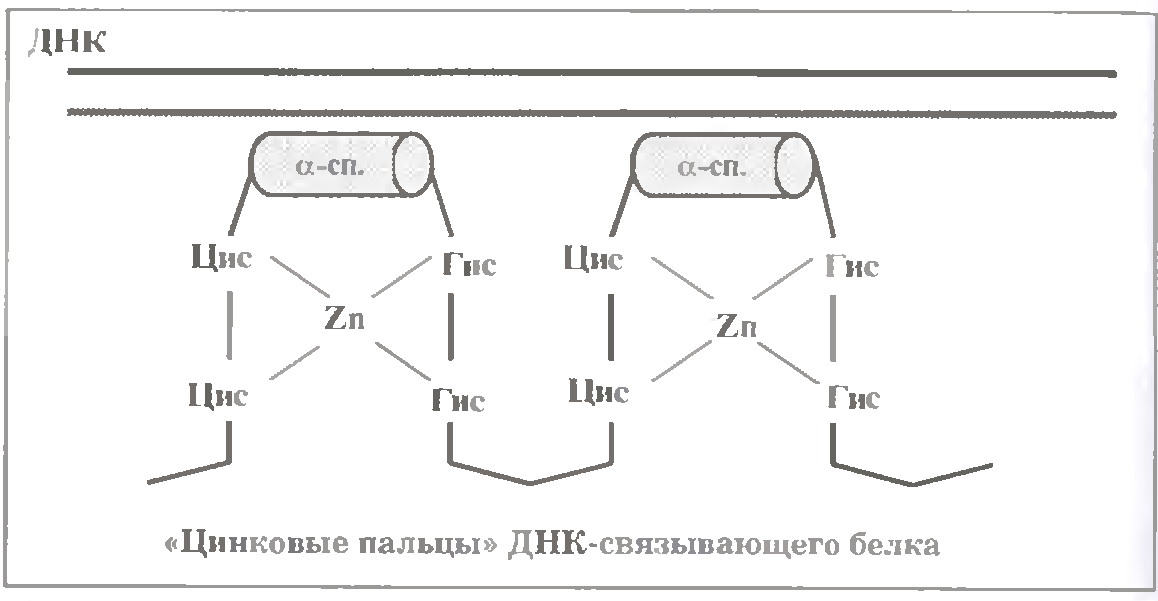
Білки цієї родини мають пальцеподібні структури (див.мал.). Кожна з останніх стабілізована атомом цинка, який утворює чотири координаційні зв'язки з двома залишками цистеїну і двома залишками гістидину пептидного ланцюга.
На зовнішній поверхні "пальця" знаходиться α-спіраль, що специфічно дізнається певну послідовність нуклеотидних пар ДНК.
Кількість "пальців" у різних білків неоднакова і сягає декількох десятків.
До цієї родини відносяться більшість регуляторних білок еукаріот. У тому числі - внутрішньоклітинні білки-рецептори стероїдних гормонів. Зв'язавши відповідний гормон, далі такий рецептор взаємнодіє "цинковими пальцями" з "своїм" енхансером у складі ДНК, що змінює транскрипційну активність певних генів.
г) Білки, що містять лейцинову"застібку".
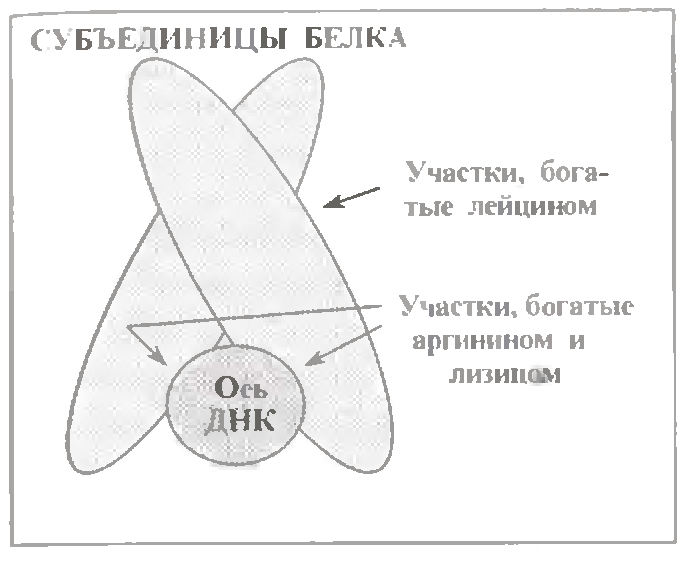
Ці білки (див.мал.) складаються з двох субодиниць, але останні можуть відрізнятися один від одного.
Головна особливість полягає в способі з'єднання субодиниць один з одним - це, в основному, гідрофобні зв'язки між залишками лейцину.
У ділянці ж зв'язування з ДНК білок багатий основними амінокислотами - аргініном і лізином. Це свідчить про те, що у взаємодії з ДНК важливе не стільки специфичне упізнавання білком певних послідовностей нуклеотидних пар (якщо таке взагалі має місце), скільки замикання іонних зв'язків з фосфатними групами ДНК.
За рахунок чого ж білок специфічно взаємодіє з потрібною ділянкою ДНК? Мабуть, усе визначається частково трьохмерною структурою цієї ділянки, а головне його білковим оточенням. Інакше кажучи, білок, що зв'язується, органічно вписується в ту просторову структуру (створювану макромолекулами), яка є в районі даного локуса ДНК. Іонні ж зв'язки просто закріплюють результат цього "вписування ".
До цієї родини відноситься більшість транскрипційнних чинників еукаріот. Тобто ті самі білки, які, як ми знаємо, взаємодіючи з ДНК, одночасно утворюють складні комплекси один з одним.
Як і гомеодоменні білки, дані білки, теж мають бактерійні "прототипи" - наприклад, Димерний білок САР (білковий активатор катаболізму).
Вищесказане підводить до наступного припущення. Чим тонша регуляція (тобто чим менше її масштаб або чим менше у регуляторного білку генів-мішеней), тим більше значення має просторова структура регульованої ділянки хромосоми.