

Вплив продуктів трансляції на розпад мРнк
Вищевикладені уявлення говорять про те, що розпад мРНК, мабуть, тісно пов'язаний з процесом трансляції. Але, можливо, грають роль і самі продукти трансляції. По принципу зворотного зв'язку вони стимулюють руйнування відповідних мРНК.
Першим цю ідею висловив в 1975 р.. В. А. Арбузов; він же намагався підтвердити її експериментальним шляхом, причому не лише для еукаріот, але і для бактерій.
Ключовим служив наступний результат: якщо трансляцію подавити антибіотиками, то середній час напівжиття мРНК зростає.
Правда, такий же ефект повинен спостерігатися і тоді, коли руйнування мРНК здійснюється 5´-РНКазой по механізму, викладеному вище. Дійсно, якщо під дією антибіотика припиняється рух рибосом по мРНК, то це повинно гальмувати також просування 5 '- РНКази, чого і означає стабілізацію мРНК.
Проте два інші факти, отриманих В. А. Арбузовим, говорили проти подібного пояснення і на користь першочергової ідеї.
а) По-перше, деякі антибіотики викликають т. з. абортивну транслокацію, при якій рибосоми продовжують просуватися по мРНК, але білок не синтезують. В цьому випадку ефект стабілізації мРНК спостерігався як і раніше.
б) По-друге, згідно з тим же джерелом, у бактерій білоксинтезуюча система пов'язана з мембранами клітини, а руйнування мРНК відбувається лише після її відділення від мембрани.
Звідси склалося наступне уявлення про механізм розпаду мРНК :
- білок (продукт трансляції) стимулює пов'язані з мембраною РНКази
- останні викликають фрагментацію мРНК цього білку
- фрагменти мРНК втрачають зв'язок з мембраною і швидко руйнуються в цитозолі розчинними нуклеазами.
Чи дійсно має місце такий механізм, поки сказати важко. З його позицій не можна, наприклад, пояснити розпад мРНК, що транслюються безпосередньо в цитозолі (т. з. мембранонезв’язані рибосомами).
Відмітимо, що тепер стали з'являтися і інші свідчення на користь початкової ідеї - про те, що білки можуть стимулювати розпад своїх мРНК. Ось два з них.
а) Розпад гістонових мРНК. Синтез гістонів активно відбувається у S-фазу синтетичного циклу, і в цей час гістонові мРНК мають термін напівжиття близько 40 хв.
Після ж накопичення гістонів (на початок G2-фази) час напівжиття їх мРНК знижується до 10 хв.
Це сам по собі дуже красномовний факт. Але не менше цікаве його конкретне пояснення.
Як неодноразово відмічалось, гістонові мРНК не мають пів(А) -фрагмента. Зате в їх 3 '- нетрансльованій області міститься шпилькова петля (мал.).
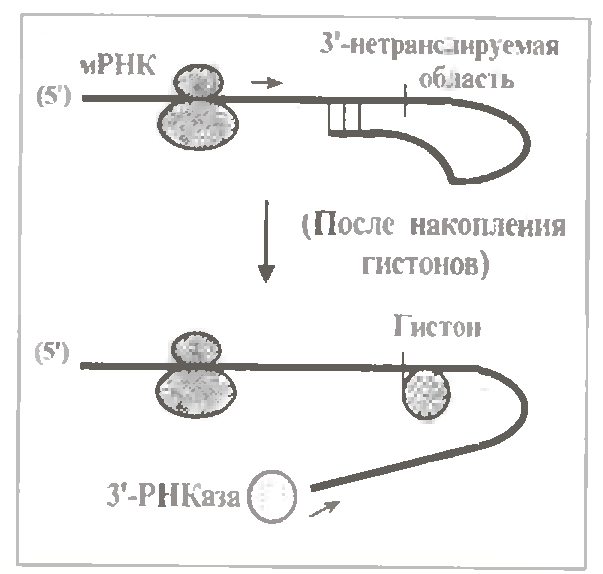
Можливо, вона так само, як петля пів(А) -фрагмента, захищає 3´-конец мРНК від РНКази. Але її особливістю є спорідненість до гістонів. Причому зв’язування гістонів призводить до прискорення деградації мРНК. Мабуть, причиною є руйнування структури петлі, що і робить 3 '- кінець доступним для РНКази.
б) Тубулінова мРНК. Розпад цієї мРНК також прискорюється продуктами її трансляції - мономерами тубуліну і навіть не до кінця синтезованим пептидом.
Механізм цього ефекту доки неясний. Але у будь-якому випадку знову йдеться про той же негативний зворотний зв'язок, який постулював В. А. Арбузов.