

Елонгація транскрипції
Етап, що йде за ініціацією, - елонгація: поступове подовження зростаючого ланцюга про-РНК до остаточного розміру.
Це відбувається у міру просування РНК-полімерази по ДНК. Відповідно, переміщається і "вічко" транскрипції, тобто ділянка локального розплітання ДНК. На транскрибованій же частині ДНК дволанцюжкова спіральна структура відновлюється відразу після відходу РНК-полімерази.
Зразкова швидкість руху ферменту і синтезу РНК - 30 нуклеотидів в секунду.
Транскрипція може супроводжуватися помилками спаровування, в результаті яких в РНК, що синтезується, включаються "неправильні" нуклеотиди. В середньому одна така помилка доводиться на 2 х 104 включених нуклеотидів. Це істотно частіше, ніж поява нерепарованих помилок реплікації (одна помилка приблизно на 1010 нуклеотидних пар).
Очевидно, менша точність транскрипції пов'язана з тим, що помилки тут мають не такі серйозні наслідки. Вони легко компенсуються завдяки утворенню на одному гені безлічі копій про-РНК.
Крім того, внаслідок виродженості генетичного коду, не кожна заміна нуклеотиду змінює зміст кодона мРНК. Можна знайти, що в 67 % помилок, що стосуються третіх положень кодонів, зміст останніх залишається тим самим.
Термінация транскрипції
Останній етап термінація, або закінчення транскрипції.
Сигналом для цього служать спеціальні ГЦ-збагачені ділянки у кінці генів. Оскільки сила взаємодії пар ГЦ досить велика, локальна денатурація таких ділянок в ДНК відбувається важче. Це уповільнює просування РНК полімерази і може служити для неї сигналом для зупинки транскрипції.
Але ще до закінчення процесу у кінці новосинтезированої РНК теж встигає з'явитися ГЦ-збагачена ділянка. Завдяки взаємодії між своїми нуклеотидами, вона утворює "шпильку". Тобто взаємодії з нуклеотидами матричного ланцюга ДНК замінюються на "внутрішньошпилькові" взаємодії. Це полегшує від'єднання РНК від ДНК.
У бактерій цьому часто сприяє і спеціальний білок - Rho-фактор. Він рухається по ДНК услід за РНК-полімеразою, наздоганяє її на ГЦ-ділянці гена і, володіючи активністю, полегшує розбіжність ланцюгів РНК і ДНК.
Конвеєрний характер процесу
Досі ми мали на увазі одну молекулу РНК-полімерази і утворення одного ланцюга про-РНК.
Але насправді через якийсь час після того, як попередня молекула РНК-полімерази, покидає промотор, просунеться по ДНК на деяку відстань, з промотором зв'язується наступна молекула ферменту і теж починає транскрипцію.
Тому на кожному гені, що транскрибується, зазвичай працюють, рухаючись один за одним, відразу декілька молекул РНК-полімерази (див.мал.).
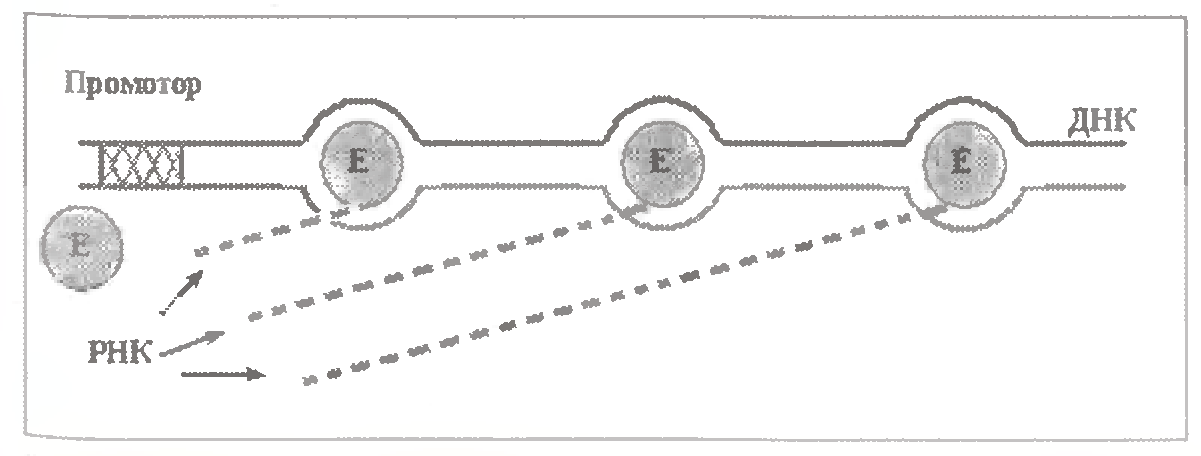
Середня відстань між ними залежить від "сили" промотора (багато в чому обумовленою транскрипційними чинниками) і концентрації РНК-полімерази. Середній порядок цієї відстані - 300-500 н. п.
Відповідно, з одним геном одночасно пов'язано декілька зростаючих ланцюгів про-РНК.
Таким чином, транскрипція гена відбувається конвейєрним способом.
Інгібітори транскрипції
Є немало речовин, що специфічно інгібують транскрипцію. Найбільш відомі з них α-аманітин і актиноміцин D.
α-Аманітин - це один з токсинів отруйних грибів (зокрема, блідої поганки). Він є цикличним пептидом, що включає ряд незвичайних амінокислот.
Цей пептид дуже міцно зв'язує РНК-полімеразу II (у еукаріот) і блокує тим самим синтез попередників матричних РНК (на стадії елонгації).
Актиноміцин D - антибіотик; точніше, це похідний природного антибіотика з Streptomytes. Він містить два ідентичних циклічних пентапептиди, сполучених з гетероциклічною системою.
Актиноміцин міцно зв'язується з ГЦ-збагаченими ділянкими ДНК. Це блокує просування РНК-полімерази - через неможливість локального розплітання ланцюгів.
Особливо чутливий до цього інгібітору синтез попередників рибосомальних РНК, оскільки їх гени дуже збагачені ГЦ-парами.