

Глава 5. Развитие глазного яблока
Сосочек Бергмайстера (Bergmeister) (рис. 5.9.2). Сосочек Бергмайстера образуется в результате накопления глиальных клеток в центре диска зрительного нерва к 9-й неделе развития. Эти клетки образуют конус, основание которого располагается на внутренней пограничной мембране, а верхушка распространяется на определенное расстояние в стекловидное тело вокруг клокетова канала. Стимулом к пролиферации этих клеток является тракция диска зрительного нерва волокнами стекловидного тела после прекращения роста первичного стекловидного тела [199]. Последующий некроз ткани приводит к образованию «физиологической чаши». При этом сосочек атрофируется. В некоторых случаях сосочек Бергмайстера сохраняется и после рождения, что не приводит к существенному изменению зрительных функций.
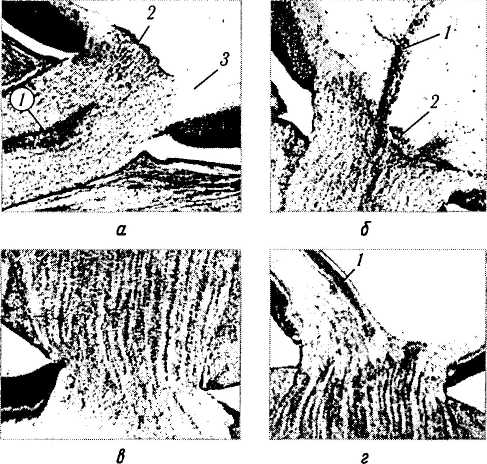
Рис. 5.9.2. Развитие зрительного нерва и диска зрительного нерва (по Duke-Elder, /963):
а — эмбрион 45 мм; б — эмбрион 67 мм; в — эмбрион 97 мм;
г — эмбрион 160 мм (1 — сосочек Бергмайстера; 2 — гиалоид-
ные сосуды; 3 — нервные волокна)
Развитие аксонов ганглиозных клеток.
На 10—12-й неделях эмбрионального развития в зрительном нерве обнаруживается до 1,9 млн аксонов ганглиозных клеток. На поперечном разрезе они имеют круглую форму, цитоплазма их бледная и содержит микротрубочки, микро-филаменты и редкие митохондрии [184, 185].
Количество аксонов быстро увеличивается, так что приблизительно к 16-й неделе эмбрионального развития зрительный нерв содержит уже 3,7 млн аксонов. К 33-й неделе 70% аксонов исчезает, и их число уменьшается до 1,1 млн.
Прогрессирующее снижение числа аксонов частично связано с дегенерацией (апоптоз)
ганглиозных клеток эмбриональной сетчатки. Определенная потеря связана и с процессами сегрегации их терминалов с нейронами наружного коленчатого тела. В последнее время показано, что процессы апоптоза нейронов сетчатки регулируют факторы, выделяемые пигментным эпителием сетчатки [239].
Рост аксонов ганглиозных клеток в направлении среднего мозга (область будущих наружных коленчатых тел) происходит благодаря наличию на их конце филоподий, между которыми располагаются как бы гофрированные мембранные образования — ламелоподии [28]. Филоподий активно взаимодействуют с окружающими структурами, проникая в ткань все глубже и глубже, в результате чего и происходит направленный рост аксонов. Именно филоподий направляют рост аксонов, определяя, таким образом, точность ретинотопической организации волокон в зрительном нерве, зрительном перекресте и зрительном пути [120]. Роль филоподий в направленном росте аксонов была доказана в эксперименте путем введения в эмбрионы, выращиваемые in vitro, цитохалазина В, который разрушает филоподий. При этом аксоны ганглиозных клеток сетчатки не достигли центров мозга [41].
Аксоны ганглиозных клеток растут со скоростью приблизительно 60—100 мкм/ч, причем скорость может изменяться в зависимости от участка пути и типа глиального окружения [78, 138]. Зоны роста аксонов чаще обнаруживаются в наружных слоях зрительного нерва, т. е. вблизи мягкой мозговой оболочки [47]. Это, по всей видимости, отражает различную скорость дифференциации ганглиозных клеток периферии и центральных областей сетчатой оболочки [46, 80, 277, 278].
В области зрительного перекреста аксоны ганглиозных клеток начинают более активно взаимодействовать с глиальными элементами. При этом ультраструктурно выявляются специализированные образования как в аксонах ганглиозных клеток, так и глиальных клеток (уплотнения мембраны и их инвагинация). Именно это взаимодействие осуществляет направление дальнейшего роста перекрещивающихся и непе-рекрещивающихся волокон зрительного нерва.
В процессе роста аксонов ганглиозных клеток, перекрещивания волокон и образования ретинотопических контактов с нейронами наружных коленчатых тел большую роль играют различные адгезивные вещества [228, 247, 248, 253]. На настоящий момент времени выявлено много подобных веществ [116, 196]. К таковым, в первую очередь, относится мембранный гли-копротеид — невральная клеточная адгезивная молекула (NCAM), обнаруженная в месте роста аксонов ганглиозных клеток сетчатки у многих животных [233, 265].
В процессе роста аксонов ганглиозных клеток сетчатки и их ретинотопической органи-
Развитие зрительного нерва
603
зации принимают участие и некоторые маркерные гены, экспрессия которых обнаруживается именно в местах перекрещивания волокон в эмбриональном периоде у многих животных. К таковым относятся TAG-1 и TAG-2. Именно эти гены активируют синтез иммуноглобулина L1, который, в свою очередь, обнаруживается в месте взаимодействия зон роста аксонов с гли-альными клетками и, по мнению ряда исследователей, предопределяет направленность роста аксонов в области перекреста [46, 48]. Если вышеприведенные факторы благоприятствуют росту аксонов, то хондроитинсульфат, наоборот, тормозит рост аксонов [34, 194].
Рост аксонов ганглиозных клеток сетчатки, их перекрест в области хиазмы и ретинотопи-ческая проекция на наружное коленчатое тело находятся под генетическим контролем. Наиболее доказано участие в этих процессах генов семейства Pax, а именно Рах-1, Рах-6, Рах-9 [272]. Подтверждением тому являются экспериментальные наблюдения, указывающие на то, что при мутации этих генов у экспериментальных животных развиваются различные аномалии глазного яблока [186]. При мутации гена Рах-2 у мышей отмечено нарушение закрытия эмбриональной щели глаза и, естественно, нарушение формирования аксонов ганглиозных клеток сетчатки, глиальных элементов [259]. Наличие мутации этого гена у человека привело к развитию колобомы и других серьезных аномалий глаза [206]. Нарушения функции ре-гуляторных генов могут привести к отсутствию перекрещивания волокон зрительного нерва в хиазме вообще. Такое состояние описано у овец [103]. Отсутствие перекрещивания волокон в хиазме у человека впервые было описано Веза-лиусом в 1543 г. Apkarian et al. [13] описали еще два таких случая.
Помимо регуляторных генов семейства Pax в регуляции роста аксонов и их перекрещивания принимают участие и другие гены, в частности гены BF-2, Dlx-2, Nkx-2 [140].
Описывая механизмы, управляющие рети-нотопической организацией волокон зрительного нерва и их перекрестом в хиазме, нельзя не остановиться и на роли нейронов, расположенных в эмбриональном периоде в области хиазмы.
Благодаря методам иммуноморфологии установлено, что маркерные гены, участвующие в регуляции роста аксонов ганглиозных клеток, обнаруживаются в нейронах промежуточного мозга, на месте которых в последующем сформируется перекрест. Эти нейроны располагаются в виде клина, острие которого направлено кпереди. Именно в этих клетках выявляется ряд веществ, непосредственно регулирующих рост аксона. Это иммуноглобулин L1, который способствует росту аксона ганглиозной клетки сетчатки, гликозилированная молекула поверхности клетки CD44, отличающаяся противо-
положным эффектом [243, 246]. Аксоны ганглиозных клеток не проникают и не пересекают те области, где в нейронах обнаруживается экспрессия указанных маркерных генов. Скорее, они как бы «отворачиваются» от них, направляясь контрлатерально и формируя крестообразную структуру хиазмы. Доказательством участия этих клеток в формировании перекреста является избирательное их повреждение при помощи антител к CD44. В результате подобного воздействия аксоны ганглиозных клеток теряют способность формировать перекрест [245]. Близкую функцию выполняют и некоторые другие маркерные гены, в частности GAP-43 [129, 244].
В регуляции перекрещивания волокон зрительного нерва в области хиазмы участвуют также глиальные элементы [115]. Правда, механизмы участия глии пока не совсем ясны. По всей видимости, вызревающие глиальные элементы тормозят рост аксонов ганглиозных клеток, направляя аксоны в область с меньшим количеством недифференцированных глиальных элементов, т. е. отклоняя их от ипсилатераль-ного распространения. Процесс регуляции роста аксонов глиальными элементами, видимо, осуществляется рядом биологически активных веществ, синтезируемых олигодендроцитами и астроцитами [114, 115, 128, 279].
Миелинизация аксонов. Миелинизация волокон зрительного нерва начинается около зрительного перекреста на седьмом месяце эмбрионального развития [6, 14, 49, 139]. Процесс миелинизации останавливается у решетчатой пластинки, что чаще всего бывает на первом месяце после рождения [266]. У новорожденных миелиновый слой вокруг аксонов ганглиозных клеток довольно тонкий и содержит большое количество холестерина. С возрастом число миелиновых слоев постепенно увеличивается.
По мере формирования зрительного нерва как анатомического образования отмечается проникновение между пучками аксонов ганглиозных клеток сетчатки соединительнотканных клеток, которые синтезируют волокна и основное вещество. В дальнейшем в этой соединительной ткани выявляются кровеносные сосуды. Параллельно с этим процессом вокруг нерва отмечается уплотнение клеток эктомезен-химы, дифференцирующейся к пятому месяцу в оболочке нерва.
Заключая этот раздел, необходимо остановиться на практическом значении сведений относительно закономерностей развития организма в целом, и глазного яблока в частности. При этом следует помнить следующее:
— во-первых, полезно знать, что патологические воздействия на эмбрион во время формирования глазного пузырька и глазного бокала могут привести к развитию разнообразных пороков развития глаза;
601