

Строение фоторецепторов
Основными светочувствительными элементами (рецепторами) являются два вида клеток: одни в виде стебелька - палочки 110-123 млн. (высота 30 мкм, толщина 2мкм), другие более короткие и более толстые - колбочки 6-7 млн. (высота 10мкм, толщина 6-7 мкм). Они распределены в сетчатке неравномерно. Центральная ямка сетчатки( fovea centralis) содержит только колбочки(до 140 тыс. на 1 мм 52 0). По направлению к периферии сетчатки их число уменьшается, а число палочек возрастает.
Каждый фоторецептор - палочка или колбочка состоит из чувствительного к действию света наружного сегмента содержащего зрительный пигмент и внутреннего сегмента, который содержит ядро и митохондрии обеспечивающие энергетические процессы в фоторецепторной клетке
{ Наружный сегмент светочувствительный участок, где световая энергия преобразуется в рецепторный потенциал Электронно-микроскопические исследования выявили, что наружный сегмент заполнен мембранными дисками , образованными плазматической мембраной . В палочках , в каждом наружном сегменте , содержится 600-1000 дисков, которые представляют собой уплощенные мембранные мешочки, уложенные как столбик монет. В колбочках мембранных дисков меньше. Это частично объясняет более высокую чувствительность палочки к свету ( палочку может возбудить всего один квант света, а для активации колбочки требуется больше сотни квантов).
Каждый диск представляет собой двойную мембрану, состоящую из двойного слоя молекул фосфолипидов, между которыми находятся молекулы белка. С молекулами белка связан ретиналь, входящий в состав зрительного пигмента родопсина.
Наружный и внутренний сегменты фоторецепторной клетки разделены мембранами, через которые проходит пучок из 16-18 тонких фибрил. Внутренний сегмент переходит в отросток, с помощью которого фоторецепторная клетка передает возбуждение через синапс на контактирующую с ней биполярную нервную клетку}
Наружные сегменты рецепторов обращены к пигментному эпителию, так что свет в начале проходит через 2 слоя нервных клеток и внутренние сегменты рецепторов, а потом достигает пигментного слоя.
Колбочки функционируют в условиях больших освещенностей - обеспечивают дневное и цветовое зрение, а палочки - отвечают за сумеречное зрение .
Видимый нами спектр электромагнитных излучений заключен между коротковолновым (длина волны от 400нм) излучением, которое мы называем фиолетовым цветом и длинноволновым излучением(длина волны до 700 нм) называемым красным цветом. В палочках находится особый пигмент- родопсин , (состоит из альдегида витамина А или ретиналя и
белка) или зрительный пурпур, максимум спектра, поглощения которого находится в области 500 нанометров. Он ресинтезируется в темноте и выцветает на свету. При недостатке витамина А нарушается сумеречное зрение -"куриная слепота".
В наружных сегментах трех типов колбочек(сине-, зелено и красно-чувствительных) содержится три типа зрительных пигментов, максимум спектров поглощения которых находится в синей (420 нм), зеленой(531 нм) и красной(558 нм) частях спектра. Красный колбочковый пигмент получил название - " йодопсин" . Структура йодонсина близка к родонсину.
{ Рассмотрим последовательность изменений:}
Молекулярная физиология фоторецепции: Внутриклеточные регистрации от колбочек и палочек животных показали, что в темноте вдоль фоторецептора течет темновой ток, выходящий из внутреннего сегмента и входящий в наружный сегмент. Освещение приводит к блокаде этого тока. Рецепторный потенциал модулирует выделение медиатора (глутамата) в синапсе фоторецептора. Было показано, что в темноте фоторецептор непрерывно выделяет медиатор, который действует деполяризующим образом на мембраны постсинаптических отростков горизонтальных и биполярных клеток.(см. Рис.1)
Палочки и колбочки обладают уникальной среди всех рецепторов электрической активностью, их рецепторные потенциалы при действии света - гиперполяризующие, потенциалы действия под их влиянием не возникают.
{ При поглощении света молекулой зрительного пигмента.(см. Рис.2) - родопсина в ней происходит мгновенная изомеризация ее хромофорной группы: 11- цис -ретиналь превращается в транс -ретиналь. Вслед за за фотоизомеризацией ретиналя происходят пространственные изменения в белковой части молекулы: она обесцвечивается и переходит в состояние метородопсина II. В результате этого молекула зрительного пигмента приобретает способность к взаимодействию с другим примембранным белком гуанозин трифосфат(ГТФ) - связывающим белком – трансдуцином (Т).
Рис.1
Рис 2.


В комплексе с метародопсином трансдуцин переходит в активное состояние и обменивает связанный с ним в темноте ганозитдифосфат(ГДФ) на (ГТФ). Трансфдуцин + ГТФ, активируют молекулу другого примеммбранного белка - фермента фосфодиэстеразы(ФДЭ). Активированная ФДЭ разрушает несколько тысяч молекул цГМФ.
В результате падает концентрация цГМФ в цитоплазме наружного сегмента рецептора. Это приводит к закрытию ионных каналов в плазматической мембране наружного сегмента, которые были открыты в темноте и через которые внутрь клетки входили Na + и Ca . Ионные каналы закрываются вследствие того, что падает концентрация цГМФ, которая держала каналы открытыми. В настоящее время выяснено, что поры в рецепторе открываются благодаря цГМФ циклическому гуанозинмонофосфату.
Механизм восстановления исходного темного состояния фоторецептора связан с повышением концентрации цГМФ. (в темновую фазу с участием алкагольдегидрогеназы+ НАДФ)
Т.о поглощение света, молекулами фотопигмента приводит к снижению проницаемости для Nа, что сопровождается гиперполяризацией, т.е. возникновению рецепторного потенциала.}
Гиперполяризационный рецепторный потенциал, возникший на мембране наружного сегмента, распространяется затем вдоль клетки до ее пресинаптического окончания и приводит к уменьшению скорости выделения медиатора - глутамата.
Фоторецепторы связаны между собой - электрическими(щелевыми) контактами. Эта связь избирательная: палочки связаны с палочками и т.д.
Эти ответы от фоторецепторов сходятся на горизонтальные клетки, которые приводят к деполяризации в соседних колбочках возникает отрицательная обратная связь, которая повышает световой контраст.
На уровне рецепторов происходит торможение и сигнал колбочки перестает отражать число поглощенных фотонов, а несет информацию о цвете, распределении и интенсивности света, падающего на сетчатку в окрестностях рецептора.
Существует 3-и типа нейронов сетчатки - биополярные, горизонтальные и амакриновые клетки. Биополярные клетки непосредственно связывают фоторецепторы с ганглиозными клетками, т.е. осуществляют передачу информации через сетчатку в вертикальном направлении. Горизонтальные и амокриновые клетки передают информацию по горизонтали.
Биполярные клетки занимают в сетчатке стратегическую позицию, поскольку все сигналы, возникающие в рецепторах поступающие к ганглиозным клеткам, должны пройти через них.
Экспериментально было доказано, что биполярные клетки имеют рецептивные поля в которых выделяют центр и переферию (Джон Даулинг- и др. Гарвардская медицинская школа).
Рецептивное поле - совокупность рецепторов, посылающих данному нейрону сигналы через один или большее число синапсов.
Размер рецептивных полей: d=10 мкм или 0,01 мм - вне центральной ямки.
В самой ямке d=2,5мкм (благодаря этому мы способны различать 2-е точки при видимом расстоянии между ними лишь 0,5 50 0 угла-2,5мкм - если сравнить то это монета в 5 копеек на расстоянии около 150 метров)
Начиная с уровня биполярных клеток нейроны зрительной системы дифференцируются на две группы, противоположным образом реагирующие на освещение и затемнение:
1 - клетки, возбуждающиеся при освещении и тормозящиеся при затемнении "on"- нейроны и
- клетки возбуждающиеся при затемнении и тормозящиеся при освещении - " off"- нейроны. Клетка с on-центром разряжается с заметно повышенной частотой.
Если слушать разряды такой клетки через громкоговоритель, то сначала вы услышите спонтанную импульсацию, отдельные случайные щелчки, а затем после включения света, возникает залп импульсов, напоминающий пулеметную очередь. Наоборот в клетках с off-реакцией (при выключении света - залп импульсов) Такое разделение сохраняется на всех уровнях зрительной системы, до коры включительно.
!!!! В пределах самой сетчатки передача информации осуществляется безимпульсным путем (распространением и транссинаптической передачей градуальных потенциалов).
В горизонтальных, биполярных и амокриновых клетках переработка сигнала происходит путем медленных изменений мембраны потенциалов(тонический ответ). ПД не генерируется.
{Чтобы понять, почему это так, следует представить себе влияние малого светлого пятна}. Рецепторы активны в темноте, а свет, вызывая гиперполяризацию уменьшает их активность. Если синапс возбуждающий, биполяр будет активироваться в темноте, а инактивироваться на свету; если же синапс тормозной, биполяр в темноте тормозится, а на свету, выключая рецептор, снимает это торможение, т.е биполярная клетка активируется. Т.о. является ли рецепторно-биполярный синапс возбуждающим или тормозным, зависит от выделяемого рецептором медиатора.
/////////////////////////////////////////////////////////// В передаче сигналов от биполярных клеток на ганглиозные участвуют горизонтальные клетки которые, передают информацию от фоторецепторов к биполярным клеткам и далее к ганглиозным.
[ Горизонтальные клетки отвечают на свет гиперполяризацией с ярко выраженной пространственной суммацией.
Горизонтальные клетки не генерируют нервных импульсов, но мембрана обладает нелинейными свойствами, обеспечивающими безимпульсное проведение сигнала без затухания.
Клетки делятся на два типа: В и С. Клетки В-типа, или яркостные, всегда отвечают гиперполяризацией вне зависимости от длины волны света. Клетки С-типа, или хроматические делятся на двух- и трехфазные. Хроматические клетки отвечают или гипер, или деполяризацией в зависимости от длины стимулирующего света.
Двухфазные клетки бывают либо красно - зеленые (деполяризуются красным светом, гиперполяризуются зеленым), либо зелено-синие (деполяризуются зеленым светом, гиперполяризуются синим). Трехфазные клетки деполяризуются зеленым светом, а синий и красный свет вызывает гиперполяризацию мембраны. Амакриновые клетки , регулируют синаптическую передачу на следующем этапе от биполяров к ганглиозным клеткам .
Дендриты амакриновых клеток разветвляются во внутреннем слое, где контактируют с отростками биполяров и дендритами ганглиозных клеток. На амакриновых клетках оканчиваются центробежные волокна, идущие из головного мозга.
Амакриновые клетки генерируют градуальные и импульсные потенциалы.(фазный характер ответа) . Эти клетки отвечают быстротекущей деполяризацией на включение и выключение света и демонстрируют слабый
пространственный антагонизм между центром и периферией.]
Выходными элементами сетчатки являются ганглиозные клетки. Ганглиозная клетка сетчатки – это первый нейрон "классического" типа в прямой цепи передачи информации от фоторецепторов к мозгу.
Различают 3 типа ганглиозных клеток. Среди ганглиозных клеток выделяют:
I. Тонические нейроны (Х-нейроны), которые возбуждаются адекватно световыми стимулами и отвечают в течении всего времени действия стимула. II. Фазические (Y-нейроны) кратковременный ответ длиной в несколько секунд.
III. W-нейроны реагирующие на движущиеся стимулы. В рецептивном поле ганглиозных клеток выделяют центр и переферию (оn - реакция), (off - реакция), (on-off реакции).
Т.е. клетки противоположным образом реагирующие на освещение и затемнение:
1- клетки, возбуждающиеся при освещении и тормозящиеся при затемнении "on"- нейроны
2- клетки возбуждающиеся при затемнении и тормозящиеся при освещении - off- нейроны.
Онтагоннистическая организация РП ганглиозных полей обусловлена наличием 2-х путей провед. в сетчатке.
Центр обладает максимальной световой чувствительностью а на краю чувствительность снижается.
Реакции на возбуждение центра связан с вертикальным путем от фоторецептора к биополярными клетками - ганглиозм.
Реакции на раздражение переферии обусловлены с горизонтальным путем проведения.
Конвергенция и диввергенция связей в сетчатке составляют основу рецептивных полей ганглиозных клеток сетчатки.
Благодаря круглой форме полей ганглиозных клеток сетчатки - происходит точечное описание сетчатого изображения, также и подкорковых ст-рах происходит взаимодействие повышение или снижение сигналов от рецепторов к мозгу.
П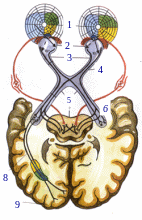 РОВОДНИКОВЫЕ
И ЦЕНТТРАЛЬНЫЕ ЗВЕНЬЯ ЗРИТЕЛЬНОГОАНАЛИЗАТОРА
РОВОДНИКОВЫЕ
И ЦЕНТТРАЛЬНЫЕ ЗВЕНЬЯ ЗРИТЕЛЬНОГОАНАЛИЗАТОРА
Схема проводящих путей зрительной системы
1-поля зрения 2 - ход лучей в глазном яблоке 3.зрительные нервы
4 - зрительный перекрест
5 - зрительный тракт
6 - наружное коленчатое тело
7 - верхние бугры
четверохолмия
8 - лучистое сияние
(пучок Грациоле)
9 - корковый центр
Зрительная информация передается в мозг через аксоны ганглиозных клеток сетчатки, которые образуют зрительный нерв. Правый и левый зрительные нервы сливаются у основания черепа и образуют Xиазму .
Нервные волокна идущие от носовых (назальных) половин обеих сетчаток, пересекаются и переходят на противоположную сторону. Волокна идущие от височных (темпоральных) половин каждой сетчатки, продолжают идти с той же стороны (ипсилатерально) объединяясь вместе с перекрещенным пучком аксонов из контрлатерального зрительного нерва и образуют зрительный тракт. В результате в правом таламусе оканчиваются волокна от правых половин каждого глаза в левом от левых.
Зрительный тракт приводит к первым зрительным станциям латеральным коленчатым телам и верхним бугоркам четверохолмия. Их аксоны проходят через заднюю часть внутренней капсулы, образуя в белом веществе полушарий мозга зрительную лучистость и заканчиваются в коре затылочной доли мозга по краям шпорной борозды. Края шпорной борозды является корковым концом зрительного анализатора.
Зрительная кора организована ретинотопически: расположение какого-либо из нейронов в этой коре соответствует локализации его рецептивного поля в сетчатке.
Рецептор - биполяр - ганглиозная клетка наружного коленчатого тела - простая клетка коры и сложная клетка.
Корковые нейроны могут быть разделены не только по рецептивным полям и по глазодоминантности и по чувствительности к движению.
Электроретинограмма
В фоторецепторах происходит трансформация световой энергии в нервное возбуждение. В рецепторах, а затем в нейронах сетчатки генерируется электрические потенциалы возникающие при увеличении или уменьшении количества света. Суммарный электрический ответ сетчатки на свет носит название электроретинограммы. Он может быть зарегистрирован от целого глаза или же непосредственно от сетчатки. Для записи электроретинограммы один электрод помещают на поверхности роговой оболочки, а другой прикладывают к коже лица вблизи глаза или на мочке.
Различают несколько типов волн: (α, β, с, d)

α - электроотрицательные колебания отражают суммацию потенциалов возникающих в фоторецепторах и горизонтальных клетках.
β- отражает изменение мембранных потенциалов глиальных клеток (мюллеровых клеток) сетчатки ионами К2+ при возбуждении биополярных и амокриновых нейронов.
с- отражает биопотенциалы пингментных клеток при «включении света»(on-эффект).
d - горизонтальных клеток фоторецепторов ( и биополярных клеток) при «выключении света» (off-эффект) (она тем больше, чем длительнее действовал свет.
Электроретинограмма хорошо отражает также такие свойства светового раздражителя как его цвет, размер, длительности действия ЭРГ на ней отраж. пот. всех клеточных элементов сетчатки (кроме ганглиозных). Этот показатель широко используется в клинике глазных болезней для диагностики и контроля лечения при различных заболеваниях сетчатки.
ЦВЕТОВОЕ ЗРЕНИЕ
Видимый нами спектр электромагнитных излучений заключен между коротковолновым (длина волны от 400 нм) излучением, которое мы называем фиолетовым цветом и длинноволновым излучением (длина волны до 700 нм) называемым красным цветом. Остальные цвета видимого спектра (синий, зеленый , желтый, оранжевый) имеют промежуточные значения длины волны. Смешение лучей всех цветов дает белый цвет. Если произвести смешение трех основных цветов - красного, зеленогои синего, то могут быть получены любые цвета.