

20.03.2013 Лекция №5
Разновидностью митоза является мейоз. Термин происходит от греч. мейо - уменьшение.
Мейоз связан с образованием гамет и наблюдается в процессе гаметогенеза. (стадия созревания)
Мейоз представляет собой особое деление, которое завершается образованием тетрады гаплоидных клеток.
Соматические клетки в рез-те мейоза формируют гаплоидные кл. В отличие от митоза, мейоз предст собой сдвоенное деление с одной полноценной интерфазой, в которой происходит репликация ДНК, и по времени она сдвинута, совпадая с профазой 1го деления.
1е мейотич деление - редукционное
2е мейотич деление - эквационное (уравнительное)
(1) предшествует полноценная интерфаза, (2) - и/ф без репликации ДНК
Биологич значение мейоза закл-ся в том, что он обеспечивает точное распределение ГМ в дочерние гаметы.
В редукционном делении происходит сложная профаза, состоящая из нескольких стадий:
- лептонема (ст. тонтих нитей)
- зигонема (сливающихся нитей)
- пахинема (толстых)
- диплонема (двойных)
- диагенез (расходящихся)
Профаза 1 - наиболее длительная стадия, и во время нее происходит изменение НМ, обеспечивающее редукцию хромосом и повышающее уровень изменчивости.
Происходит конъюгация гомологичных хромосом с образованием особых структур - бивалентов (синаптонемные комплексы): гомологичн хромосомы подходят близко друг к другу, выравниваются аллельными генами и образуют структуру, представляющую собой диаду по количеству хромосом и тетраду по количеству хроматид.
После конъюгации происходит кроссинговер - обмен участками гомологичных хромосом, что приводит к возникновению новой комбинации сочетания генов (сх.1)
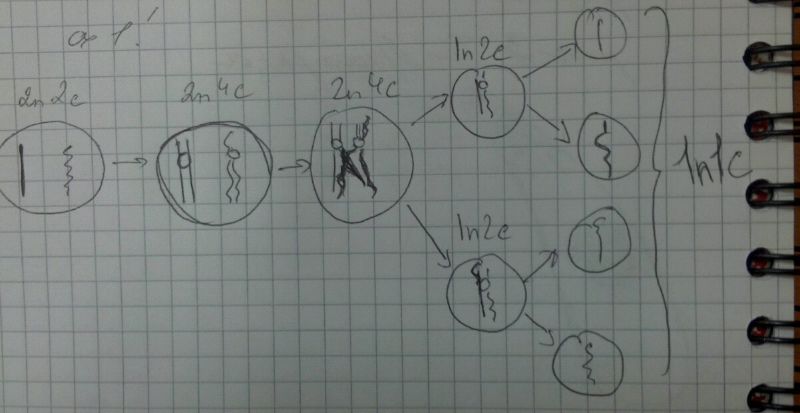
Второе деление называется эквационным. В результате этого деления образуются клетки, идентичные материнским
В анафазе редукционного деления к полюсам, а затем к дочерним клеткам расходятся гомологичные хромосомы от каждого бивалента
Во втором делении количество хромосом не изменяется, а количество ДНК еще раз уменьшается в 2 раза, и образуются 4 гаплоидные клетки
В результате мейоза количество хромосом уменьшается в 2 раза, а количество ДНК - в 4.
В анафазе второго деления расходятся сестринские хроматиды.
За счет кроссинговера, происходящего в профазе 1 мейоза, происходит рекомбинация генов, что повышает уровень изменчивости и, следовательно, мейоз представляет собой средство генетической адаптации. Это открывает возможности для отбора и существования в условиях переменной среды.
Цитологические основы мейоза связаны со свободным, случайным комбинированием хромосом вначале в диадах, а затем в тетрадах. В результате мейоза образуются стабильно молодые клетки, которые способны дать начало развитию любого организма.
Генетические аспекты оплодотворения.
В оплодотворении принимают участие 2 разнокачественные гаметы от 2 родительских особей и, в рез-те слияния этих гамет возникает особая диплоидная клетка - зигота, с генетическими потенциями обоих родителей. Оплодотворение осущетсвляет 2 функции:
1. половая (связана с комбинированием генов 2 родителей) - обеспечивает передачу генов от родителей потомкам.
2. репродуктивная (непосредственно создание новых организмов) - обеспечивает инициацию цитопл реакций, позволяющих продолжить развитие.
В генетическом отношении мужские и женские гаметы представляют собой уникальные структуры. Они содержат гаплоидный набор ГМ и характеризуются особенностями строения ядра и цитоплазма.
В ядрах спермиев ДНК сильно уплотняется, а в ядрах женских гамет наблюдается одно или несколько ядрышек. Ядра гамет генетически эквивалентны, но имеют функциональные различия, которые обусловлены разными модификациями ДНК.
После оплодотворения ядра гамет закономерно изменяются и преобразуются в мужской и женский пронуклеусы. Последние начинают перемещаться навстречу друг другу, и во время перемещения происходит репликация ДНК. Пронуклеусы сближаются и при их контакте ядерные оболочки разрушаются. Затем происходит конденсация хроматина с образованием видимых хромосом.
У большинства животных хромосомы расп-ся на общем митотическом веретене первого деления дробления.У большинства млекопитающих истинно диплоидное ядро появляется не у зиготы, а у двухклеточного зародыша (после 1го деления дробления).
Гаметы существенно отличаются и по содержанию цтп элементов. Женская гамета содержит большие запасы РНК, белка, морфогенетических факторов и рибосом - все это обеспечивает развитие по т.н. женскому типу на ранних этапах развития.
Также гаметы отличаются по размерам. Женская значительно крупнее мужской, неподвижна; мужская - маленькая, осн массу составляет ядро с ГМ, а на стпдии дифференциации происходит перемещение ГМ, и он распред-ся след. образом:
головка сперматозоида - сод. ГМ
тело - сод. осн массу цтп с органеллами (пластич и энергет комплекс)
хвостик - обесп механич движение.
Оплодотворение обеспечивает материальную непрерывность между поколениями и сочетанием потоков признаков родителей.
Сочетание родит признаков во время оплодотворения является результатом не простого суммирования генов, а следствием взаимодействия генов, что обеспечивает активное развитие.
Для оплодотворения 1 женской гаметы необходима 1 мужская гамета. Это моноспермия.
У ряда животных (насекомые, рыбы, земноводные, рептилии, некот млекопит) - нормальна полиспермия - проникновения неск спермиев. Оплодтворение все равно моносперное. ост сперматозоиды выполняют трофическую и регуляторную функцию.
В практике осеменения животных используют факультативную полиспермию, притом семенная ж-ть часто смешанная, что способствует повышению многоплодия и улучшению хозяйственных признаков.
При оплодотворении происходит случайная встреча разных гамет, что приводит к тому, что среди особей 1 вида практически невозможно появление двух генетически одинаковых организмов
Оплодотворение обеспечивает сохранения постоянства числа хромосом вида и повышает изменчивость потомства. Яйцеклетки оплодотворяются спермтозоидами только своего вида, что возможно обусловлено генетическим сродством гамет одного вида друг к другу. --- между видами существует избирательность оплодотворения, причем у высших животных в результате естественного отбора возникли особенности (морфолог, физиолог, биохим и поведенч), обеспечивающие эту избирательность (разные сезоны спаривания, различное строение и форма половых органов). В генетическом отношении даже при равнм количестве хромосом невозможно обеспечить объединение хромосом в гомологичные пары.
Между разными видами избирательность оплодотворения выражена достаточно резко. Внутри вида избирательность отсутствует, что является положительным фактором эволюции,. т.к способствует возрастанию комбинативной изменчивости. Внутри вида сущетсвует только одно самоограничение, и оно связано с самооплодотворением.
Избирательность не связана с генетическим составом гамет, т.к в это время ДНК находится в спирализованном замкнутом состоянии.
Процесс оплодотворения происходит при определенных условиях.
1. определенный рН в половых путях самки
2. наличие маркерных белков-АГ в гаметах
в некоторых случаях при оплодотворении обнаруживается несовместимость гамет, что приводит к бесплодию и ухудшению качества потомства.
Существуют организмы, которые способны развиваться и без оплодотворения - партеногенез. Причем для некоторых видов это норма (тни, дафнии, долгоносики). При партеногенезе развитие может быть успешным в условиях достаточчной гетерогенности я/к. Мб гаплоидным и диплоидным.
Гиногенез. развитие нередуцированной я/к стимулируется проникновением в нее спермия. Слияния пронуклеусов не происходит. Пример: серебристый карась.
Еще разновидность: андрогенез. Развитие я/к связывют с мужским ядром. До оплодотв ядро я/к погибает --- ложное оплодотворение, развитие за счет ядра мужской гаметы.
кроме того, существуют особые формы полового процесса: полиандрия и полигения.
Полиандрия (-спермия) - с участием лишних сперматозоидов. Генетически обнаруживается наличием неск мужских пронуклеусов.
Полигения - в результате оплодотворения наряду со с/з принимают участие 1 или неск женских пронуклеусов и цитогенетически это обн-ся в соединении одного мужского и 2 и более жен пронуклеусов.