

Ряд дятлоподібні - Piciformes
Включає майже 400 видів птахів (маса від 6 до 300 г), ведучих переважно деревний спосіб життя. Їжу збирають у кронах і на стовбурах, рідше - на землі. Моногами. Гніздяться в дуплах - природними або видовбаних самими птахами, деякі риють гніздові нори. Пташенята лупляться сліпими й голими. У гнізді залишаються, поки повністю не оперяться й не стануть здатними до польоту. Ряд розділяють на два підряда.
У підряді примітивні дятлоподібні - Galbulae включають 170 видів, у яких ознаки деревної спеціалізації не настільки чіткі й різноманітні, як у дійсних дятлів. Поширені в лісах і чагарникових заростях тропічної зони. Якамарові - Galbulidae, пуховкові – Bucconidae, ідоові - Capitonidae - дрібні птахи (маса менш 100 г), харчуються комахами, плодами й насіннями. У лісах Африки й Південно-Східної Азії живуть зовні схожі на синиць 12 видів медоуказчиків - Indicatoridae. Харчуються різними комахами. Розшукують гнізда ос і бджіл, поїдаючи личинок, мед і соти. Здатні переварювати віск. Всі види - типові гніздові паразити, підкладають яйця в гнізда дятлів і бородаток. У лісах Центральної й Південної Америки живуть 40 видів туканових - Ramphastidae (маса 100-300 г). Дуже довгий і зовні надзвичайно потужний дзьоб (див. мал. 37) легкий (кістки губчаті, ріг тонкий). Їдять плоди; ловлять комах, грабують гнізда дрібних птахів.
Підряд дійсні дятли - Pici поєднує близько 200 видів птахів з типовим виглядом дятлів (див. мал. 37), розповсюджених у помірних і тропічних широтах обох півкуль (немає в Австралії). Дзьоб прямий, міцний, долотоподібний. Два пальці спрямовані вперед, два - назад; кігті міцні, сильно вигнуті. Тонкий й довгий язик може висуватися на довжину дзьоба; щетинки на його кінці дозволяють витягати комах з вузьких щілин і ходів. Липка слина, що покриває язик, полегшує схоплювання здобичі. При пересуванні по стовбурах дятли чіпляються за нерівності кори гострими кігтями й опираються на твердий хвіст. Різноманітних комах збирають на стовбурах і гілках, рідше - на землі, і видовбують з-під кори й з деревини. Грабують гнізда дрібних птахів. Їдять насіння; узимку видовбують насіння із шишок хвойних дерев. Щорічно в загниваючому дереві роблять гніздове дупло; деякі види займають природні дупла. Види, що потрапили в безлісні ландшафти, роблять гніздові нори в обривах й у термітниках.
У СРСР гніздиться 13 видів тільки з підряду дійсних дятлів. Найбільш широко розповсюджений великий строкатий дятел — Dendrocopos major. Самий великий з наших дятлів — чорний дятел, або желна, — Dryocopus martius (маса близько 300 г) населяє высокостовбурні ліси. У змішаних і листяних лісах майже повсюдно живе вертишийка — Jynx torquilla, що відрізняється від інших дятлів більш слабким дзьобом, прямо обрізаним хвостом з м'яких кермових. По вертикальних стовбурах лазити не може.
Ряд Горобцеподібні - Passeriformes
Величезний ряд, що поєднує близько 5000 видів птахів (тобто приблизно 60% всіх нині живучих птахів), розповсюджених по всіх континентах, крім Антарктики, що мають різні розміри, різний зовнішній вигляд й екологічні особливості. Особливо різноманітні горобцеві тропічних лісів. Є твариноядні, рослинноядні й всеядні види. Звичайно моногами, але в деяких видів є полігамія. Будують гнізда у гілках, ущелинах скель, у дуплах, на землі, у норах гризунів і т.д. Пташенята лупляться сліпими, голими або слабко опушеними; у гнізді залишаються, поки не досягнуть розмірів дорослих і не оперяться. Велика кількість ряду змушує виділяти в ньому 50—70 сімейств, що груперуються у чотири підряди.
Підряд рогодзьоби — Eurylaimi включає 14 видів лісових птахів, розповсюджених в Африці й Південно-Східній Азії. По ряду особливостей схожі на ракшеподібних, що свідчить про філогенетичну близькість цих рядів.
Підряд кричащи горобці, або тиранові — Clamatores (Tyranni) поєднує близько 1000 видів, розповсюджених у тропіках обох півкуль. Особливо різноманітні й численні в Південній Америці. Нижня гортань має слабко розвинену мускулатуру, але, незважаючи на це, частина видів прекрасно співає, а деякі здатні й до звуконаслідування.
У підряді примітивні горобці — Menurae - всього 4 види, що зустрічаються в деяких районах Австралії. Птахи ліри (p. Меnurd) зовні нагадують фазанів (маса 500—800 г). Живуть у лісах з густим підліском. Швидко бігають, злітають неохоче. Токуя, самець розправляє хвіст так, що він стає схожим на ліру. Пологамори. Будує гніздо, насиджує єдине яйце й вигодовує пташеняти тільки самка.
Підряд співочі горобці — Passeres (Oscines) включає близько 4000 видів. Поширені повсюдно. Лише в Південній Америці по різноманіттю видів і по чисельності уступають кричащим горобцям. 330 видів горобців, що зустрічаються в СРСР відносяться тільки до цього підряду. Розміри (маса від 4 г до 1,5 кг), зовнішній вигляд, звички досить різноманітні. У нижній гортані добре розвинена 5-7 пара голосових м'язів, а нижні кільця трахеї зростаються в кістковий барабан - резонатор. Нижче згадується лише частина сімейств цього підряду.
Близько 70 видів родини жайворонкові — Alaudidae живуть у відкритих ландшафтах. Задній палець закінчується довгим прямим кігтем. Годуються на землі безхребетними й насінням. Гнізда на землі. Самці часто співають у повітрі. Родина ластівкові - Hirundinidae включає близько 80 видів прекрасно літаючих гострокрилих пташок. Дзьоб маленький, але розріз рота великий (суглоб на рівні ока). По землі ходять мало. Здобич - різних комах - ловлять у повітрі або на льоту скльовують із гілок, каменів і т.п. Практично космополіти. Гніздяться по долинах рік, узліссям лісів, багато видів - синантропи (гніздяться в поселеннях людини). Частина видів будує ліпні гнізда із грудочок бруду, скріплюючи їхньою липкою слиною; інші риють нори в обривах або гніздяться в дуплах, ущелинах і т.п.
У родині воронів - Corvidae близько 100 досить великих видів (маса 50 г - 1,5 кг). Великодзьобі, великоголові кремезні птахи. Населяють найрізноманітніші місцеперебування. Гнізда на деревах, у заломах очеретів, у тріщинах обривів, у деяких — у дуплах. Багато видів всеїдні. У СРСР гніздиться 15 видів, у тому числі ворон — Corvus corax (маса близько 1,5 кг), ворона — С. corone, грак — С. frugilegus, сорока — P. pica, сойка — Garrulus glandarius та ін.
У сирих лісах Нової Гвінеї й Австралії зустрічається близько 20 видів бесідкових птахів - Ptilonorhynchidae, по зовнішньому вигляді нагадують дроздів. Перед початком розмноження самець розчищає площадку діаметром 1-3 м і прикрашає її камінчиками, квітами, ягодами; зів'ялі квіти і ягоди заміняються свіжими. В інших видів на площадці будується бесідка із трави, стінки якої теж прикрашаються. З появою самки починаються танці, у яких беруть участь обоє партнера, і спарювання. Потім самка в гіллі будує гніздо, відкладає 2 яйця, насиджує й годує пташенят.
Родина райські птахи - Paradisaeidae поєднує близько 40 видів. Це рухливі птахи, розмірами від синиці до галки, населяють густі ліси Нової Гвінеї, кілька видів зустрічається в Австралії. Самці забарвленні яскраво й мають пір'я, що утворюють чуби, комірці, пучки на боках або крилах і т.п. Самки забарвленні тускло. Характерне групове токовище: 10- 20 самців, зібравшись у кроні дерева, приймають різноманітні пози, міняють положення прикрашаючого пір'я, злітають із лементами й знову сідають і т.п. Самки підлітають до місця токовища й відбувається спарювання. Пари не утворяться. Вся турбота про потомство падає тільки на самку. Посилено переслідувалися (яскраве пір'я використалися для прикрас). Тому чисельність сильно зменшилася, а ряд видів повністю винищені.
Близько 60 видів дрібних рухливих лісових птахів становлять родину синицеві — Paridae; у нас зустрічається 13 видів (див. мал. 37). Дуплогніздники; лише ремези (Remiz sp.) з рослинних волокон і пуху в'ють мішкоподібні гнізда з бічним входом, підвішуючи їх на тонких гілках. Кладки до 10 і навіть до 16 яєць. Насиджує частіше самка, а самець її годує, пташенят вигодовують обоє. Годуються різними комахами і їхніми кладками, їдять ягоди й насіння. Легко залучаються в культурні ландшафти розввішуванням штучних гнізд. Дуже корисні як винищувачі різноманітних шкідливих комах.
Поползневі — Sittidae — короткохвості птахи із сильним прямим дзьобом — населяють ліси й гори Євразії й Північної Америки. Гніздяться в дуплах або тріщинах скель; зменшують вхід в укриття, замазуючи його брудом. Легко лазають по стовбурах і скелях. Харчуються безхребетними, насіннями, ягодами. Корисні винищуванням шкідливих комах. У нас широко розповсюджений звичайний повзик — Sitta europaea (див. мал. 37). К піщуховим — Certhiidae ставляться рухливі пташки з довгим, тонким, вигнутим донизу дзьобом, що населяють ліси й гори всіх континентів, крім Південної Америки. Легко лазають по стовбурах і скелям, вибираючи із тріщин різних безхребетних. У нас звичайна піщуха — Certhia familiaris (див. мал. 37).
Родина оляпкові — Cinclidae включає всього 5 видів кремезних короткохвостих птахів (маса 50—80 г), що живуть по берегах швидких річок Європи, Азії, Північної Африки й Америки. Харчуються безхребетними і їхніми личинками, зрідка ловлять дрібну рибу. Їжу збирають, бігаючи по берегах або пірнаючи (єдині серед горобиних пірнаючі птахи). Присівши на воду птах тримає крила так, що плином течії її прибиває до дна. Пірнає на глибину 1—1,5 м і пробігає по дну, збираючи корм, 10— 20 м, під водою залишається до 15—20 с. Коли птах складає крила, його, як пробку, викидає на поверхню, і він злітає. Пірнати у воду з повільною течією оляпки не можуть. У СРСР у передгірних і гірських районах більш звичайна звичайна оляпка — С. cinclus.
Близько 60 видів крапивникових — Troglodytidae живе в лісах, чагарникових і кактусових заростях степів і пустель, по скелястих узбережжях Північної й Південної Америки. Це комахоїдні кремезні пташки з тонким і гострим дзьобом, злегка вигнутим донизу. Звичайний кропивник — T. troglodytes живе не тільки в Північній Америці, але й на більшій частині території Євразії (у тому числі й у СРСР).
Родина дроздові — Turdidae поєднує близько 300 видів птахів різноманітних розмірів (маса 10—200 г) і зовнішнього вигляду. Заселяють всі наземні ландшафти. Харчуються безхребетними, насіннями, ягодами. У СРСР зустрічається близько 50 видів. Це різні дрозди p. Turdus, солов'ї — Luscinia, горихвістки — Phoenicurus, каменки -— Oenanthe, карбівки — Saxicola та ін. Багато видів — прекрасні співаки. Більше 400 видів дрібних (вага 6—40 г), рухливих птахів відносяться до сімейства славкові — Sylviidae. Більшість видів зустрічається тільки в східній півкулі, заселяючи тут всі ландшафти. Багато видів — прекрасні співаки. Харчуються різними безхребетними, при нагоді їдять ягоди. Перелітні. У СРСР зустрічається 50 видів: різні славки — Sylvia, очеретники — Acrocephalus й ін.
Близько 400 видів дрібних (маса 8-30 г) птахів східної півкулі, що населяють ліси, чагарникові зарості й культурні ланшафты, відносяься до родини мухоловки - Muscicapidae. Частина видів будує гніздо в розвилці гілок, інші гніздяться в дуплах, охоче займають штучні гнізда. Перелітні. Зазвичай птах сидить на гілці, злетівши, ловить комах, і знову сідає; при нечисленності літаючих комах збирають їх на гілках, листках і на землі. Іноді їдять ягоди. У СРСР зустрічається 14 видів.
Родина трясогузки — Motacillidae поєднує близько 50 видів струнких дрібних птахів (маса 20—30 г) з характерним виглядом ковзана або трясогузки, що населяють відкриті ландшафти або узлісся. У СРСР зустрічається 15 видів. Майже повсюдно гніздиться біла трясогузка — Motacilla alba.
Близько 70 видів сорокопутових — Laniidae поширені в Африці, Євразії, Північній Америці. Дзьоб міцний, закінчується гачком. Хвіст подовжений. Тримаються по узліссям, по чагарниках у степах, пустелях і горах. Поїдають великих комах, ловлять ящірок і гризунів, грабують гнізда дрібних птахів. Велику здобич наколюють на сучок і потім розривають дзьобом; на сучки біля гнізда наколюють і надлишок здобичі. У СРСР зустрічається 11 видів. Широко розповсюджений сорокопуд жулан — Lanius cristatus.
Близько 100 видів утворюють родину шпакові — Sturnidae. Заселяють відкриті ландшафти й узлісся східної півкулі. Широко розповсюджений по території СРСР звичайний шпак — Sturmis vulgaris успішно акліматизований у Північній Америці й на багатьох островах. Гнізда в укриттях - дуплах, під каменями, у щілинах будівель і т.п. Годуються на землі, у гілках дерев і кущів, поїдаючи безхребетних, ягоди, насіння, проростки рослин. Корисні знищенням масових видів шкідливих комах. Під час кочівель і перельотів місцями можуть заподіювати збиток, роздзьобуючи вишні, виноград й інші плоди. У нас гніздяться 6 видів.
У тропіках східної півкулі поширені близько 250 видів дрібних (маса 6—20 г), яскраво забарвленних птахів з довгими тонкими, часто вигнутими дзьобами, яких відносять до двох родин — медососні — Meliphagidae і нектарники — Nectariniidae. Харчуються нектаром і комахами, яких викльовують із квітів. Виконують роль запильників.
Близько 300 видів ткачикових — Ploceidae широко поширені в східній півкулі; більш різноманітні в тропіках, особливо в Африці. Більшість видів зерноядні, але їдять і ягоди, і різноманітних безхребетних; пташенят вигодовують звичайно комахами. Часто гніздяться групами й навіть колоніями. У СРСР зустрічається 11 видів. Найбільше широко поширені горобці: будинковий — Passer domesticus (акліматизований в Америці) і польовий — P. montanus.
Майже 500 видів включають у родину в'юркові — Fringillidae. Поширені вони широко (немає в Австралії), заселяють найрізноманітніші ландшафти — відкриті й лісові — від тундр до гір. Зазвичайно самці забарвленні яскравіше самок. Багато видів — гарні співаки. Поза періодом розмноження тримаються зграйками. Переважно поїдають багато безхребетних і сім'я. Годуються в кронах, на траві й на землі. Пташенят вигодовують комахами. У СРСР зустрічається 70 видів. Більше інших звичайний зяблик — Fringitla coelebs, клести — Loxia sp., щіголь — С. carduelis, чиж — S. spinus, снігур — P. pyrrhula, різноманітні вівсянки p. Emberiza (у СРСР їх гніздиться 24 виду) та ін.
Особливості організації птахів
Форма тіла. Пристосування до польоту обумовило відносну одноманітність форми тіла. Тулуб компактний, більш-менш округлий. Голова невелика, шия довга й рухлива. Передні кінцівки - крила - у спокійному стані складені й притиснуті до боків тіла. Оперення забезпечує обтічність тіла. Видові варіації в розмірах і формі дзьоба й голови, довжині шиї, довжині й формі крил і хвоста, довжині задніх кінцівок і формі їхніх пальців забезпечують при збереженні загальної одноманітності пристосування до різних типів руху й харчування.
Розміри птахів варіюють у невеликих межах; можливість польоту обмежує зростання розмірів. Маса великих літаючих птахів не перевищує 14—16 кг (лебеді, грифи, дрохви) при розмаху крил до 3—4 м (пелікани, альбатроси). Самі дрібні із птахів — деякі колібрі з максимальною масою 1,6—2 гр. Втрата здатності до польоту часто приводить до збільшення розмірів: маса великих пінгвінів досягає 40 кг, казуарів й африканських страусів — 80—100 кг. Деякі з вимерлих птахів, що не літали, досягали маси 300-400 кг (эпіорніси, моа).
Шкіра і її похідні. Шкіра птахів тонка, суха, практично позбавлена шкірних залоз. Поверхневі шари клітин эпідермального шару стають роговими. Сполучнотканинний шар шкіри підрозділяється на тонку, але досить щільну власне шкіру, у якій проходять кровоносні судини, укріплені закінчення контурного пір'я й розташовані пучки гладких мускульних волокон, що міняють положення пір'я, і підшкіряну клітковину - пухкий шар, що безпосередньо прилягає до тулубної мускулатури; у ньому відкладаються запаси жиру. Єдина шкіряна залоза - куприкова - лежить на хвостових хребцях. Вона виробляє жироподібний секрет, що виділяється через протоки, коли птахи надавлюють на залозу дзьобом. Птахи змазують цим секретом пір'я, що сприяє збереженню їхньої еластичності та підвищує водовідштовхувальні властивості оперення. Жирові виділення куприкової залози під впливом сонячного світла перетворюються у вітамін D, який птахи заковтують при очищенні пера. Куприкова залоза добре розвинена в більшості птахів, ведучих як водний, так і наземний спосіб життя (гусеподібні, куреподібні й ін.); лише в деяких наземних і водних птахів вона розвинена слабко (баклани, чаплі) або відсутній (страусоподібні, дрохви, деякі папуги).
Розростання зроговівшого эпідермального шару шкіри утворюють роговий покрив дзьоба - рамфотеку. Рогові лусочки рептильного типу покривають пальці, цівку, а іноді й частину гомілки. Останні фаланги пальців ніг покриті роговими кігтями. У самців деяких птахів (наприклад, уфазанових) на цівці утворився кістковий виріст, покритий гострим роговим чохлом, - шпора. Специфічний для птахів пір'яний покрив - теж рогові утворення эпідермального шару шкіри.
Основний
тип пера — контурне перо. Воно складається
з міцного й пружнього рогового стовбура,
з боків якого розташовані м'які зовнішнє
й в нутрішнє
опахала (мал. 38). Частина стовбура, до
якої прикріплюються опахала, називають
стрижнем; він у поперечному розрізі
має чотиригранну форму. Нижня, позбавлених
опахал частина стовбура називається
очином і має округлий розріз; підстава
очина занурена в шкіру і зміцнюється
в пір'яній сумці. Кожне опахало, що
відходять від бічних сторін стрижня,
утворене подовженими роговими пластинками
— борідками першого порядку, від яких
у свою чергу відходять численні більш
тонкі борідки другого порядку (бородочки)
з розташованими на них дрібними
крючочками. Крючочки, зчіплюючись із
сусідніми борідками, утворюють пружну
платівку опахала. Якщо, наприклад при
ударі, крючочки розійдуться й опахало
«розірветься», птах, дзьобом поправляючи
перо, змусить крючочки знову зчепитися,
і структура пера відновиться. Звичайно
в самій нижній частині пера борідки
більш тонкі й пухнаті, без гачків; це
пухова частина опахала, функція якої
— утримувати в шкірі шар повітря. В
осілих птахів зимове перо має більш
розвинену пухову частину, ніж літнє. У
частини птахів від черевної поверхні
стрижня на рівні нижнього краю опахала
відходить додатковий стрижень (мал.
38, 8),
м'які
борідки якого теж не мають крючечків.
Розвиток пухових додаткових стрижнів
збільшує теплоізоляційні якості пера.
нутрішнє
опахала (мал. 38). Частина стовбура, до
якої прикріплюються опахала, називають
стрижнем; він у поперечному розрізі
має чотиригранну форму. Нижня, позбавлених
опахал частина стовбура називається
очином і має округлий розріз; підстава
очина занурена в шкіру і зміцнюється
в пір'яній сумці. Кожне опахало, що
відходять від бічних сторін стрижня,
утворене подовженими роговими пластинками
— борідками першого порядку, від яких
у свою чергу відходять численні більш
тонкі борідки другого порядку (бородочки)
з розташованими на них дрібними
крючочками. Крючочки, зчіплюючись із
сусідніми борідками, утворюють пружну
платівку опахала. Якщо, наприклад при
ударі, крючочки розійдуться й опахало
«розірветься», птах, дзьобом поправляючи
перо, змусить крючочки знову зчепитися,
і структура пера відновиться. Звичайно
в самій нижній частині пера борідки
більш тонкі й пухнаті, без гачків; це
пухова частина опахала, функція якої
— утримувати в шкірі шар повітря. В
осілих птахів зимове перо має більш
розвинену пухову частину, ніж літнє. У
частини птахів від черевної поверхні
стрижня на рівні нижнього краю опахала
відходить додатковий стрижень (мал.
38, 8),
м'які
борідки якого теж не мають крючечків.
Розвиток пухових додаткових стрижнів
збільшує теплоізоляційні якості пера.
Різноманітне забарвлення птахів забезпечується як нагромадженням у клітинах пера в період його формування пігментів, так і мікроскопічними особливостями структури пера. Основні типи пігментів — меланіни й ліпохроми. Накопичення в рогових клітинах глибки й зернят меланіну обумовлюють чорне, буре й сіре забарвлення Ліпохроми у вигляді жирових крапель або пластів залягають у роговій речовині, забезпечуючи червоне, жовте й зелене забарвлення. Сполучення в ділянці пера різних пігментів ускладнює забарвлення. Білий колір обумовлений наповненою повітрям безбарвною роговою масою пера. Властивий перу багатьох птахів мінливий по відтінку металевий блиск, створюється завдяки інтерференції світла в зовнішніх оболонках рогових клітин. Пігменти, що забарвлюють перо, підвищують його механічну міцність. Забарвлення оперення має різноманітне значення: полегшує зустріч особин одного виду, запобігає міжвидовим зіткненням, часто робить птаха малопомітним в його звичайному місцеперебуванні й т.п.

Рис. 39. Розташування птерилій (1) і аптерій (2) на шкірі
голуба
Контурні пір'я, що покривають все тіло птаха, укріплені в шкірі на особливих полях - птериліях (мал. 39), розділених аптеріями - ділянками шкіри, на яких пір'я не ростуть. Таке черепицеподібне розташування пір'я, що налягає одне на одне, дозволяє закрити все тіло мінімальною кількістю пір'я. Лише в деяких птахів, що переважно не літають (страусоподібні, пінгвіни), пір'я рівномірно розподілені по всій поверхні шкіри. Довгі й особливо міцні пір'я утворюють площину крила; їх називають маховими. Першорядні махові прикріплюються до заднього краю скелету кисті, другорядні - до заднєверхньої сторони ліктьової кістки. Махове пір'я розташовано так, що зовнішнє опахало покриває лише частину більш широкого внутрішнього опахала сусіднього пера. При опусканні крила пір'я утворюють суцільну площину, що давить на повітря (мал.40). При підніманні крила махові трохи повертаються й між ними утворяться щілини, через які проходить повітря. Довгі й міцні пір'я, що утворюють площину хвоста, називають кермовими.
Під контурним пір'ям лежить пухове пір'я; у них тонкий стрижень, а борідки не несуть крючечків, тому не утворюється зчеплене опахало.
РРРис 40. Схема положення махового пір'я і направлення опору повітря (стрілки) при опусканні крила (/) і при його підйомі (//) (по Гессе)
Пух — це пухове перо, у якого різко вкорочений стрижень і довгі, сильно опушені борідки відходять пучком від кінця очина. Пух і пухові пір'я або рівномірно покривають все тіло (гусеподібні, веслоногі й ін.), або розташовані тільки по аптеріях (чаплі, сови, багато горобиних). Пух і пухові пір'я забезпечують теплоізоляцію. У багатьох птахів по всьому тілу розташоване нитковидне пір'я з тонким стовбуром і рідкими короткими борідками. Вони служать датчиками, що сигналізують про токовища повітря під пір'яним покривом. У кутах рота в багатьох птахів розташовані щетинки; це пір'я із пружним стрижнем, що втратили борідки. Вони виконують тактильну функцію, а в частини видів, що ловлять на льоту дрібну здобич (дрімлюги, ластівки, стрижі), збільшують розміри ротового отвору.
Розвиток пера. У результаті розростання клітин эпідерміса й кутиса на шкірі утворюється горбок, що поступово розростається у вигляді спрямованого назад виросту, підстава якого заглиблюється в шкіру, утворюючи піхва пера. Сполучнотканинна частина виросту перетворюється в пронизаний кровоносними судинами сосочок зростаючого пера. Одночасно эпідермальний шар, що розростається, диференціює на тонкий роговий чохлик, що закриває зростаюче перо, а з посилено ділячихся й поступово ороговіваючих підлягаючих клітин формується стрижень із борідками, що відходять від нього. У міру росту пера навколишній його роговий чехлик поступово злущується, а борідки розпрямляються, утворюючи опахало. Повністю виросле перо - мертве утворення, що утримується припасованими до очина стінками піхви й мускульних пучків коріуму. Їхнє скорочення дозволяє міняти положення пера (разпушувать або, навпаки, притискати оперення). Сосочок засихає; від нього в очині пера залишається лише дужку з плівкою (див. мал. 38, 4).
Згодом пір'я зношуються й вицвітають, погіршуються їх механічні й теплоізоляційні властивості. Тому відбувається періодична зміна пера - лінька. Повна лінька зі зміною всього пір'я звичайно проходять наприкінці літа, після завершення розмноження. Звичайно при цьому здатності птахів до польоту помітно не погіршуються. Однак у деяких груп (гусеподібні, пастушки, журавлі й ін.) зміна контурного пір'я по тілу йде поступово, а махові й кермові з їх криючими випадають одночасно, і птах втрачає здатності до польоту (дрібні качки — приблизно на 20 днів, лебеді — майже на 1,5 місяця); птахи, що линяють, тримаються у важкодоступних місцях. У частини видів у році буває не одна, а дві линьки. Друга звичайно йде ранньої навесні й захоплює не все оперення: кермові і махові не міняються. Наявність двох линянь забезпечує можливість сезонної зміни забарвлення і якості оперення. Так, у тетерячих під час осіннього линяння розвивається більш довге покривне пір'я із сильніше розвинутою пуховою частиною опахала й з довгим, пухнатим додатковим стрижнем, що істотно підвищує теплоізоляційні якості зимового вбрання в порівнянні з літнім. В осілих дрібних птахів у зимовому вбранні буває більше пір'я, чим у літньому, що теж забезпечує кращу теплоізоляцію взимку: наприклад, у чижів в зимовому вбранні 2100-2400 пір'їн, а в літньому - близько 1500.
Рухова система й основні типи руху. Рухи птахів різноманітні: ходьба, стрибки, біг, лазання, плавання, пірнання, політ. Вони забезпечуються як змінами опорно-м'язової системи, так і перетвореннями інших систем органів, що здійснюють координацію рухів й орієнтування в просторі, що створюють необхідні енергетичні резерви. Своєрідна особливість кістяка птахів — добре виражена пневматичність кісток. Плоскі кістки мають губчату будову, зберігаючи більшу міцність при невеликій товщині. Трубчасті кістки теж тонкостінні, а порожнини усередині них заповнені частково повітрям, частково — кістковим мозком. Ці особливості забезпечують підвищену міцність окремих кісток і помітно їх полегшують. Потрібно, звернути увагу на те, що загальна маса кістяка становить 8—18% маси тіла птахів — приблизно стільки ж, скільки в ссавців, у яких кістки товще, а повітряні порожнини в них відсутні. Це пояснюється тим, що в птахів полегшення кісток дозволило різко збільшити їхню довжину (довжина кістяка ноги, а особливо крила в кілька разів перевищує довжину тулуба), помітно не підвищуючи загальної маси кістяка.
Як й в інших вищих хребетних, кістяк птахів підрозділяється на осьовий скелет і пов'язану з ним грудну клітину, череп, скелет кінцівок й їхніх поясів (мал. 42).
Осьовий скелет — хребетний стовп підрозділяється на п'ять відділів: шийний, грудний, поперековий, крижовий і хвостовий. Число шийних хребців змінюється — від 11 до 23—25 (лебеді). Як й у плазунів, перший хребець — атлас, або атлант, — має форму кісткового кільця, а другий — эпістрофєй — зчленовується з ним зубовидним відростком; це забезпечує рухливість голови щодо шиї. Інші шийні хребці птахів гетероцельного типу,
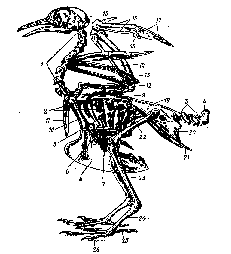
Рис. 42. Кістяк голуба: / — шийні хребці, 2 — грудні хребці, 3 — хвостові хребці, 4 — пнгостнль, 5 — спинна частина ребра із крючковндиым відростком, 6 — черевна частина ребра, 7 — грудина, 8 — кіль грудини, 9 — лопатка, 10 — коракоид, // — вилочка, 12 — плече, 13 — променева кістка, 14 — ліктьова кістка, 15 — пястно-запястная кістка (пряжка), 16 — перший палець, 17 — другий палець, 18 — третій палець, 19 — подвздошная кістка, 20 — сіднична кістка, 21 — лобкова кістка, 22 — стегно, 23 — гомілка, 24 — цівка, 25 -перший палець, 26 — четвертий палець
довге тіло кожного хребця попереду й позаду має сідлоподібну поверхню (у сагиттальном розрізі хребці опистоцельны, а у фронтальном — процельны). Зчленування таких хребців забезпечує їхню значну рухливість відносно один одного в горизонтальній і вертикальній площинах. Міцність з'єднань хребців підсилюється наявністю суглобних відростків на підставах верхніх дуг, що утворять між собою ковзні суглоби. Шийні ребра птахів рудиментарний зростаються із шийними хребцями, образуя канал, по якому проходять хребетна артерія й шийний симпатичний нерв. Тільки останні одне-два шийних ребра зчленовуються із шийними хребцями рухливо, однак до грудини вони не доходять. Особливості шийних хребців разом зі складно диференційованими шийними м'язами дозволяють птахам вільно повертати голову на 180°, а деяким (сови, папуги) і на 270°. Це уможливлює складні й швидкі руху головою при схоплюванні рухливого видобутку, чищенню оперення, будівлі гнізда; у польоті дозволяє, згинаючи або розгинаючи шию, у певних межах міняти положення центра ваги, полегшує орієнтування й т.п.
Грудних хребців у птахів 3—10. Вони зростаються один з одним, утворюючи спинну кістку, і дуже тугим суглобом з'єднуються зі складним хрестцем. Завдяки цьому тулубний відділ осьового кістяка стає нерухомим, що важливо при польоті (коливання тулуба не заважають координації літальних рухів). До грудних хребців рухливо причленовуються ребра. Кожне ребро складається із двох відділів — спинного й черевного, що рухливо зчленовуються один з одним утворюючи кут, вершина спрямована назад (мал. 42, 5—6). Верхній кінець спинного відділу ребра рухливо причленовується до поперечного відростка й тіла грудного хребця, а нижній кінець черевного відділу - до краю грудини. Рухливе зчленування спинного й черевного відділів ребер між собою й рухливе їхнє з'єднання із хребетним стовпом і грудиною поряд з розвинутою реберною мускулатурою забезпечують зміну обсягу порожнини тіла. Це один з механізмів інтенсифікації подиху. Міцність грудної клітини підсилюється крючковидними відростками, укріпленими на спинних відділах і налягають на наступне ребро. Велика грудина має вигляд тонкої широкої й довгої пластинки, на якій у всіх птахів (крім страусоподібних) розташований високий кіль грудини (crista sterni). Велики розміри грудини і її кіля забезпечують місце для прикріплення потужних м'язів, що рухають крило.
Всі поперекові, крижові і частина хвостових хребців нерухомо зростаються один з одним у монолітну кістку — складний хрестець (synsacrum). Усього в нього входять 10—22 хребця, границі між якими не видні. Зі складним хрестцем нерухомо зростаються кості тазового пояса. Це забезпечує нерухомість тулубного відділу й створює міцну опору для задніх кінцівок. Число вільних хвостових хребців не перевищує 5—9. Останні 4—8 хвостових хребців зливаються в сплощену з боків куприкову кістку pygostyle (мал. 42, 4), до якої віялом прикріплюються основа кермового пір'я. Укорочення хвостового відділу й утворення пігостиля забезпечує міцну опору хвосту при збереженні його рухливості. Це важливо, тому що хвіст не тільки виконує функцію додаткової несучої площини, але бере участь й у керуванні польотом (як гальмо й кермо).
Череп птахів схожий на череп рептилій і може бути віднесений до діапсидного типу зі скороченою верхньою дугою. Череп тропібазальный (очниці розташовані поперед головного мозку), утворений тонкими губчатими костями, границі між якими чітко видні лише в молодих птахів. Це пов'язане з тим, що з'єднання за допомогою швів неможливо через невелику товщину кісток. Тому череп відносно легкий. Своєрідна в порівнянні із плазунами і його форма: різко збільшений об'єм мозкової коробки, очниці більші, щелепи позбавлені зубів (у сучасних птахів) і формують дзьоб. Зсув великого потиличного отвору й потиличного мищєлка на дно черепа збільшує рухливість голови щодо шиї й тулуба.
Великий потиличний отвір оточений чотирма потиличними кістками: основний (basioccipitale) (мал. 43, 1), двома бічними (occipi- tale latеrale) і верхньої (supraoccipitale). Основна й бічні потиличні кістки утворюють єдиний (як й у плазунів) потиличний мищєлок, що зчленовується з першим шийним хребцем. Три вушні кістки, що оточують слухову капсулу, зливаються із прилеглими костями й між собою. У порожнині середнього вуха є лише одна слуховая кісточка — стремінце. Боки й дах мозкової коробки утворюють парні покривні кістки: лускаті (мал. 43; 21), тім'яні, лобові й бічні клиноподібні (laterosphenoideum) (мал. 43, 9). Дно черепа утворює покривна основна клиноподібна кістка, котру за- криває покривна основна скронева кістка (basitemporale), і клювовидний відросток парасфеноїда (rostrum parasphenoidei) (мал. 43, 7, 8). У його переднього кінця лежить сошник, по краях якого знаходяться хоани.
Верхня частина дзьоба — наддзьоб — утворений сильно розрозшимися, що злилися, передщелепними кістками (мал. 43, 17). Гребінь дзьоба, укріплений носовими кістками, з'єднується з лобовими костями й передньою стінкою очниці, утвореною розрозшимися середньою нюховою кісткою (mesethmoideum). Складову лише задньої частини наддзьоба верхньощелепні кістки відростками зливаються з піднебінними кістками (мал. 43, 28). До заднєзовнішнього краю верхнєщелепної кістки приростає тонка кісткова поперечина, що складається із двох кісток, що злилися, - скулової і квадратно-скулової. Це типова нижня дуга діапсидного черепа, що обмежує знизу очницю й скроневу яму. Квадратно-скулова кістка зчленовується із квадратною кісткою, нижній кінець якої утворює суглобну поверхню для зчленування з нижньою щелепою, а подовжений верхній кінець суглобом прикріплюється до лускатої й передньовушної кісток. Піднебінні кістки кінцями налягають на дзьобовидний відросток парасфеноїда й суглобом з'єднуються з парними крилоподібними кістками, які у свою чергу суглобом пов'язані із квадратними кістками відповідної сторони.
Така будова кісткового піднебіння має важливе значення для властивого більшості птахів кінетизма (рухливості) наддзьба. При зкорочені м'язів, що з'єднують спрямований уперед очноямковий відросток квадратної кістки зі стінкою очниці, нижній кінець квадратної кістки зміщається вперед і зрушає як піднебінні так і крилоподібні кістки (їхнє з'єднання один з одним може сковзати по дзьобовидному відростку), так і квадратно-вилична й вилична.

Рис. 43. Череп молодого голуба. А — збоку; Б — знизу; В — зверху:
1 —. основна потилична кістка, 2 — бічна потилична кістка, 3 — потиличний виросток, 4 — великий потиличний отвір, 5 — верхня потилична кістка, 6 — зовнішній слуховой прохід, 7 — основна скронева кістка, 8 — дзьобовидний відросток, 9 — бічна клиноподібна кістка, 10 — ококлиноподібна кістка, 11 — міжочна перегородка, 12 — середня нюхова кістка, 13 — тім'яна кістка, 14 — лобова кістка, 15 — носова кістка, 16 — предлобная кістка, 17 — передщелепна кістка, 18 — верхньощелепна кістка, 19 — вилична кістка, 20 — квадратно-вилична кістка, 21 — луската кістка, 22 — квадратна кістка, 23 — сошник, 24 — крилоподібна кістка, 25 — сочленова кістка, 26 — зубна кістка, 27 — кутова кістка, 28 — піднебінна кістка

1
— гнучка зона наддзьоба, 2
—
з'єднання наддзьоба з піднебінним
апаратом, 3
—
щелепний суглоб, 4
—
квадратно-черепний суглоб
Тиск по цих кісткових містках передається на основу наддзьоба й завдяки перегину кісток в області «перенісся» (мал. 44, /) вершина наддзьоба зрушується догори. У зоні перегину наддзьоба кістки дуже тонкі, а в деяких видів (гусаки й ін.) тут утворюється суглоб. При скороченні м'язів, що з'єднують череп з нижньою щелепою, вершина дзьоба зрушується донизу. Рухливість кісткового піднебіння в сполученні зі складно диференційованими жувальними м'язами забезпечують різноманітні, тонко диференційовані рухи дзьоба при ловлі здобичі, чищенню оперення, будівлі гнізд. Імовірно, рухливість шиї й пристосування дзьоба до різноманітних рухів сприяли перетворенню передніх кінцівок у крила, тому що заміняли деякі з виконуваних ними другорядних функцій (допомога в захопленні їжі, чищення тіла й ін.).
Нижня частина дзьоба — піддзьоб або нижня щелепа — утворюється злиттям ряду кісток, з яких найбіль великі зубна (див. мал. 43, 26), зочленовна й кутова. Щелепний суглоб формують зочленовна й квадратна кістки. Рух наддзьоба й піддзьоба дуже чітко координовані завдяки диференційованій системі жувальних м'язів. Під'язичний апарат складається з подовженого тіла, що підтримує основу язика, і довгих ріжків. У деяких птахів, наприклад дятлів, дуже довгі ріжки обгинають весь череп. При скороченні під'язичної мускулатури ріжки сковзають по сполучнотканному ложу і язик висувається з ротової порожнини майже на довжину дзьоба (мал. 45).
1—
язик, 2
—
ріжки під'язичного апарата аются
в загальну п'ясно-зап'ясну кістку
(carpometacarpus), або пряжку (див. мал. 42, 15).
Різко
редукується кістяк пальців: добре
розвинені тільки дві фаланги другого
пальця (див. мал. 42,17),
що
продовжують
вісь пряжки. Від першого і третього
пальця зберігається лише по одній
короткій фаланзі (див. мал. 42, 16,
18). Першорядні
махові прикріплюються до пряжки й до
фаланг другого пальця. До фаланги
першого пальця прикріплюється кілька
пір'їн «крильця».
аются
в загальну п'ясно-зап'ясну кістку
(carpometacarpus), або пряжку (див. мал. 42, 15).
Різко
редукується кістяк пальців: добре
розвинені тільки дві фаланги другого
пальця (див. мал. 42,17),
що
продовжують
вісь пряжки. Від першого і третього
пальця зберігається лише по одній
короткій фаланзі (див. мал. 42, 16,
18). Першорядні
махові прикріплюються до пряжки й до
фаланг другого пальця. До фаланги
першого пальця прикріплюється кілька
пір'їн «крильця».
Перетворення кисті (утворення пряжки, редукція пальців, мала рухливість суглоба) забезпечують міцну опору першорядним маховим, що випробовує в польоті найбільші навантаження. Характер поверхонь всіх суглобів такий, що забезпечує легку рухливість лише в площині крила; можливість обертових рухів різко обмежена. Це попереджує вивертання крила, дозволяє птахові без зусиль міняти площу крила в польоті й складати його в спокої. З'єднуючий кистьовий згин із плечовим суглобом складка шкіри - літальна перетинка (patagium) - утворює еластичний передній край крила, що згладжує ліктьовий згин і запобігає утворенню тут завихрень повітря. Характерна для кожного виду форма крила визначається довжиною кістякових елементів і другорядних і першорядних махових.
Пристосування до польоту чітко виражені й у поясі передніх кінцівок. Потужні коракоїди розширеними нижніми кінцями міцно з'єднуються малорухомими суглобами з переднім кінцем грудини (див. мал. 42, 10). Вузькі й довгі лопатки зростаються з вільними кінцями коракоїдів, утворюючи глибоку суглобну западину для голівки плеча. Міцність кісток плечового пояса і їхнє міцне з'єднання із грудиною забезпечує крилам опору в польоті. Подовження коракоїдів збільшує площу прикріплення м'язів крила й виносить уперед, на рівень шийних хребців, плечовий суглоб; це дозволяє укладати крило збоку тулуба в спокої й вигідно аеродинамічно, тому що в польоті центр ваги птаха знаходиться на лінії, що з'єднує центри площ крил (забезпечується стійкість). Ключиці зростаються у вилочку (furcula) (див. рис 42, 11), розташовану між вільними кінцями коракоїдів і виконують роль амортизатора, що зм'якшує поштовхи при взмахах крила.
Задні кінцівки й тазовий пояс зазнали перетворення, пов'язаних з тим, що при русі по суші на них переноситься вся вага тіла. Кістяк задньої кінцівки утворений потужними трубчастими костями. Загальна довжина ноги навіть в «коротконогих» видів перевищує довжину тулуба. Проксимальний кінець стегна (див. мал. 42, 22) закінчується округлою голівкою, що зчленовується з тазом, а на дистальному кінці рельєфні поверхні утворюють із костями гомілки колінний суглоб. Його зміцнює лежача в мускульному сухожиллі колінна чашечка. Основний елемент гомілки - кістковий комплекс, якому можна назвати тібіотарзус (tibiotarsus), тому що до добре розвинутої великої гомілкової кістки (tibia) приростає, утворюючи її дистальиый кінець, верхній ряд кісточок передплюсни. Мала гомілкова кістка (fibula) сильно скорочена й приростає до верхньої частини зовнішньої поверхні великої гомілкової кістки. Її редукція пов'язана з тим, що в більшості птахів всі елементи кінцівки рухаються в одній площині, обертові рухи в дистальній частині кінцівки обмежені.
Дистальний (нижній) ряд кісток передплюсни (tarsus) і всієї кістки плюсни (metatarsus) зливаються в єдину кістку — цівку, або плюсно-передплюсну (tarsometatarsus) (див. мал. 42, 24); з'являється додатковий важіль, що збільшує довжину кроку. Тому що рухливий суглоб розташовується між двома рядами елементів передплюсни (між костями, що злилися з великою гомілковою кісткою, і елементами, що ввійшли до складу цівки), те його називають інтертарзальным. До дистального кінця цівки прикріплюються фаланги пальців.
Як й у всіх наземних хребетних, тазовий пояс птахів утворений трьома парами кістками, що зростаються. Широка й довга клубова кістка зростається зі складним хрестцем. До її зовнішнього краю приростає сіднична кістка (див. мал. 42), з якою зростається гілковидна лобкова кістка. Всі три кістки беруть участь в утворенні вертлужної западини, в яку входить, утворюючи тазостегновий суглоб, голівка стегна. Лобкова й сіднична кістки в птахів не зростаються один з одним по середній лінії тіла; такий таз називають відкритим. Він дає можливість відкладати великі яйця й, сприяє інтенсифікації подиху, не обмежуючи рухливості черевної стінки в тазовій області.
Велика поверхня таза і його міцне з'єднання з осьовим кістяком забезпечують опір заднім кінцівкам і створюють можливість для прикріплення потужних ножних м'язів. Довгі міцні кістки кінцівок, різкий рельєф їхніх суглобних поверхонь поряд з добре розвинутою і диференційованою ножною мускулатурою забезпечують інтенсивний рух у різноманітних умовах.
Мускулатура птахів диференційована сильніше, а її відносна маса більша, ніж у плазунів. Це пов'язане з більшою рухливістю птахів і розмаїтістю їхніх рухів. Компактність тіла, обумовлена вимогами аеродинаміки, у значній мірі досягається тим, що найбільш потужні м'язи, що здійснюють рухи кінцівок, розташовані на тулубі, а їхнього сухожилля йдуть до кінцівок. Сильний розвиток зв'язок зміцнює з'єднання окремих елементів кістяка. Дуже складна мускулатура шиї, що забезпечує високу рухливість голови, що важливо й при схоплюванні здобичі, і при орієнтуванні, і в польоті.
З м'язів передньої кінцівки в першу чергу заслуговують згадування дві. Підключичний м'яз (musculus subclavius) прикріплюється на коракоїді, тілі й гребені грудини, а її сухожилля закінчується на голівці плеча; скорочення цього м'яза піднімає крило. Над нею лежить, прикріплюючись до грудини і її кілю, до коракоїду й коракоїдно-ключичної зв'язки, більший грудний м'яз (m. pectoralis major), що опускає в польоті крило; її сухожилля прикріплюється теж до голівки плеча. Обидва великих грудних м'яза становлять від 10 до 25% від загальної маси птаха й перевищують масу підключичних м'язів в 3-20 раз. Особливо великі ці м'язи в птахів, що літають стрімким, маневреним польотом. Крім цього, роботою крила в польоті керують ще кілька десятків більш дрібних м'язів, що розташовуються на тулубі, плечі й передпліччі.
Рух задніх кінцівок здійснюють більше 30 м'язів. Більші з них широкими основами прикріплюються до кісток таза, дрібні м'язи розташовані на стегні й гомілці. Розташований на задній поверхні гомілки глибокий згинач пальців (m.flexor digitorum perforans) утворює сухожилля, що проходить по задній стороні интертарзального суглоба й цівки, а потім розділяється на чотири гілки й, що закінчується на нижній поверхні кінцевих фаланг пальців. Поверхня кінцевих сухожиль і дно широких сполучнотканиних піхв, по яких вони рухаються, мають поперечну ребристість. Коли птах сідає на гілку й стискає пальці, під тиском її маси сухожилля притискаються до стінки піхв, і їхні реберця зчіплюються: пальці залишаються в стислому стані й при розслабленні м’язу. Щоб розійшовся цей «автоматичний замок», потрібно скорочення м'язів - разгиначів пальців. Це дає можливість птахам спати, сидячи на гіллі, з розслабленою мускулатурою.
Дихальні рухи грудної клітини здійснюються за допомогою міжреберних й інших м'язів стінок тіла. Декількома м'язами здійснюється рух хвоста. У порівнянні із плазунами, у птахів краще розвинена підшкірна мускулатура, що дозволяє міняти положення пір'я на більших ділянках тіла. Дрібні мускульні пучки коріуму міняють положення окремого пір'я.
Для птахів характерне накопичення в м'язах міоглобіну, що дозволяє створювати резервний запас кисню, що утилізується у період інтенсивної роботи. Найвища концентрація міоглобіну відзначена у великому грудному м'язі, мускулатурі мускульного шлунка й серця. Концентрація м'язового гемоглобіну вище в птахів, що літають активним польотом, у пірнаючих птахів і птахів високогір’я. У всіх випадках концентрація гемоглобіну в крові вище, чим у м'язах.
Лазання й стрибки в гілках за допомогою задніх і передніх кінцівок і використання крил для польоту - це основні способи руху примітивних древніх птахів. І зараз більшість видів у тоум або іншому ступені пов'язані із кронами дерев і кущів. У гілках птахи звичайно пересуваються стрибками, іноді допомагаючи одиночними змахами крил. Звичайний для них тип будови лапи - три пальці вперед, один назад - дозволяє міцно охоплювати гілки. Подальша спеціалізація до деревного способу життя часто супроводжується зміною будови лапи - два пальці спрямовані вперед, два назад, що, імовірно, ще більш полегшує втримання на гілках. Прекрасно лазаючі в кронах папуги використають для хапання й потужний дзьоб. За рахунок різкого посилення мускулатури ніг, розвитку сильних пальців з гострими пазурами багато хто, особливо дрібні, види птахів освоїли лазання по вертикальних стовбурах (повзики, пищухи й ін.) і по скелях. Сильні лапи з потужними пальцями й гострими пазурами й міцний хвіст, служать додатковою опорою, дозволяють дятлам не тільки лазити по вертикальних стовбурах, але, утримуючись на одному місці, довбати кору й деревину.
Багато видів деревних птахів збирають корм і на землі. Дрібні види при цьому, як й у гілках, стрибають (горобці й ін.), а інші ходять і бігають, поперемінно переставляючи ноги (біла трясогузка, грак, ворона й ін.). Пристосування до наземного способу життя часто супроводжується вкороченням пальців, особливо спрямованого назад (іноді він редукується), подовженням цівки. Кращі бігуни серед птахів – ті, що втратили здатність до польоту, страусоподібні птахи. Трипалий південноамериканський нанду й двопалий африканський страус можуть бігти зі швидкістю більше 50 км/год. У мешканців боліт і берегів водойм (чаплі, пастушки, кулики й ін.) подовження цівки й гомілки дозволяє бродити по мілководдю, не змочуючи пір’я, а довгі тонкі пальці не дають провалюватися на грузлому ґрунті. У глухаря, рябчика й інших тетерячих під час осіннього линяння з боків пальців виростають рогові шипики, що збільшують площу опори й зменшують провалювання в сніг, а в білих куріпок на пальцях виростає довге пір'я, що перетворюють лапу в лижу - снігоступ. Багато плаваючих птахів добре ходять по суші (чайки й ін.); у деяких гарних нирців ноги далеко віднесені назад і по суші вони пересуватися майже не можуть (гагари, поганки). Мало й погано ходять деякі добре літаючі види (ластівки, щурки, стрижі).
Плавання й пірнання. Завдяки малій щільності (оперення, повітряні мішки) можуть сідати на воду й злітати з неї й наземні птахи, наприклад дрозди. Здатність до дійсного плавання, коли птах перебуває на воді, годуючись і відпочиваючи, виробилася в представників різних рядів. У гарних плавців тіло трохи сплющене в спинно-черевному напрямку, що надає більшу стійкість на воді, сильно розвинена пневматичність кістяка, оперення більш щільне, чим у наземних птахів. При плаванні ноги трохи ставляться назад, а сила гребка збільшується тим, що пальці з'єднані плавальною перетинкою (всі чотири - у пеликаноподібних і тільки три, спрямовані вперед, - у трубконосих, гагар, чайок, гусеподібних) або кожен палець обгортає шкіряста облямівка (поганки, лисухи, кулики плавунчики).
Пірнання для птахів затруднено через їхню низьку щільність. Незважаючи на це деякі види успішно добувають їжу в товщі води. Пірнаючих птахів можна розділити на дві групи.
До першого ставляться види, що пірнають із пікірування, коли за рахунок інерції птах поринає у воду на невелику глибину (на 1-2 довжини тіла), схоплюючи в цей момент здобич, а потім викидається, як пробка, назовні. Так пірнають як добре плаваючі птахи (деякі пеликаноподібні, трубконосі, чайки), так і птахи, що не вміють плавати, наприклад скопа й орлан білохвіст із соколоподібних, зимородки з ракшеподібних й ін. Пристосувань до пірнання в них мало: небагато щільніше оперення, висока швидкість пікірування й т.п.
Друга група - дійсні пірнаючі птахи, пірнаючи, поринають на більшу глибину (деякі види - на кілька десятків метрів) і активно рухаються в товщі води, відшукуючи й переслідуючи здобич. У таких видів виражені пристосування до пірнання. Зменшується пневматичність кісток. Підвищується щільність (до 0,7-0,8); притискаючи оперення й довільно зменшуючи обсяг повітряних мішків у момент пірнання, птахи здатні підвищити щільність майже до 1,0. Укорочення крил поліпшує обтичність тіла у воді, але супроводжується погіршенням польоту. Стегновий суглоб відходить назад. Тому на суші тіло здобуває більш-менш вертикальне положення (баклани, чистикові, особливо пінгвіни), хода робиться незграбною. Рух у товщі води відбувається за допомогою гребних рухів ніг, що роблять різкі поштовхи назад. Чистикові птахи в товщі води працюють напіврозправленими крилами («підводний політ»:), а витягнуті назад ноги з розправленими пальцями виконують лише роль руля. У пінгвінів при плаванні на поверхні води й при пірнанні функцію рушія виконують перетворені в ласти крила, а ноги працюють лише як рулі.
Тривалість пірнання не перевищує 2-3 хв, лише в деяких видів (пінгвіни) - до 5-7 хв. У пірнаючих птахів вище резерви кисню: більше гемоглобіну в крові й у м'язах. У момент занурення зменшується число серцевих скорочень, а кров'ю продовжує інтенсивно забезпечуватися лише головний мозок і деякі внутрішні органи (кишечник, нирки). Працюючі м'язи використають кисень, зв'язаний м'язовим гемоглобіном, і переходять на анаеробні процеси (гліколіз), про що свідчить різке підвищення концентрації молочної кислоти після пірнання в м'язах й у крові. У товщі води звичайно рухаються зі швидкістю 1-2 м/с; пінгвіни здатні розвивати швидкість до 8-10 м/с (до 36 км/год).
Найбільш специфічна форма руху птахів, що визначила основні риси організації цього класу, — політ. Аеродинаміка польоту птахів складна й дотепер відома лише загалом. Зв'язано це з тим, що в польоті міняється положення махових і площа крила, з різною швидкістю й під різними кутами до обрію рухаються кистьова й основна частини крила й т.п. Грубо схематично фізичну основу польоту можна характеризувати так. Крило завжди зверху більш-менш опукло, а знизу ввігнуте, передній край крила більш товстий (тут лежить кістяк, м'язи й кілька шарів пір'я), задній — тонкий й еластичний (утворений лише вершинами махових). Обтікаючи верхню опуклу поверхню крила, зустрічний потік повітря прискорює рух, і над крилом утворюється область зниженого тиску. Створюється піднімальна сила, піднімаюча крило нагору. Коли в польоті птах опускає крило, одночасно протікають два процеси. Першорядні махові кінцевою частиною крила, що відгинаються під тиском повітря, створюють ефект пропелера, у результаті чого виникає тягова сила, що штовхає крило (і птаха) уперед. Одночасно повітря обтікає основну частину крила (область другорядних махових) і тут створюється, за рахунок різниці тисків над крилом і під крилом, піднімальна сила, що переборює силу ваги птаха. При підйомі крила нагору махові трохи повертаються, пропускаючи повітря (див. мал. 40), і тому підйом відбувається з меншим зусиллям. Рухаюча нагору й назад вершина крила створює деяку додаткову силу тяги, а основна частина крила як і раніше створює піднімальну силу. Піднімальна сила виникає й при обтіканні повітрям тіла й хвоста птаха, що летить.
Такий політ, коли птах ритмічно піднімає й опускає крила, називається маховим. Змінюючи площу крила і його нахил («кут атаки»), варіюючи частоту змахів, птах змінює величину тяги й піднімальної сили, міняючи тим самим швидкість і висоту польоту. Розходження в розмірах і формі тіла, розмірах і формі крил і хвоста, в інтенсивності й амплітуді змахів крила визначають властивий кожному виду характер польоту. Повільний, зі спокійними й рідкими змахами крил політ чаплі відмінний від стрімкого й маневреного польоту ластівок і стрижів і від швидкого, але прямолінійного польоту качок. Але всі ці птахи літають маховим польотом. Одна з форм махового польоту - тріпотливий політ, коли птах, посилено працюючи крилами, на короткий час «повисає» у повітрі на одному місці. Так роблять чайки, крачки, дрібні хижаки, що виглядають здобич. Подібним чином «повисають» у повітрі біля квітки колібрі; при цьому крило робить 50-80 змахів у секунду.
Д ругий
тип
польоту
— ширяючий; птах з розпростертими
практично нерухомими крильми
переміщається, використовуючи енергію
потоків повітря. Розрізняють статичне
й
динамічне
ширяння. Статичне ширяння можливо над
материками, де стійкі висхідні струми
повітря виникають на стиках ландшафтів
(ліс
і поле й
т.п.)
або при
обтіканні
повітрям перешкод — обривів, гірських
вершин. Для птахів, що використають
стійкі струми повітря, характерні більш
широкі, закруглені крила з розбіжними
на кінцях вершинами першорядних махових.
Такий політ використають хижі птахи,
лелеки, пелікани. Широкими колами птахи
поступово набирають висоту й
потім
кружляють, виглядаючи здобич, або,
плануючи із втратою висоти, переміщаються
в потрібному напрямку (мал. 46). Динамічне
ширяння властиве морським птахам
(альбатроси, буревісники, чайки), що
мають довгі, але вузькі, із загостреною
вершиною крила. Використовуючи завихрення
повітря над хвилями або різною швидкістю
повітряних потоків, птах по вітрі планує
вниз, набираючи швидкість, і біля води,
де швидкість вітру вповільнена тертям
об воду, повертається проти вітру й
злітає вгору, де повітря рухається
швидше (мал. 47). Так птах може ширяти
годинниками, виглядаючи здобич і
добуваючи її з пікірування. При
відсутності вітру ці птахи ширяти не
можуть.
ругий
тип
польоту
— ширяючий; птах з розпростертими
практично нерухомими крильми
переміщається, використовуючи енергію
потоків повітря. Розрізняють статичне
й
динамічне
ширяння. Статичне ширяння можливо над
материками, де стійкі висхідні струми
повітря виникають на стиках ландшафтів
(ліс
і поле й
т.п.)
або при
обтіканні
повітрям перешкод — обривів, гірських
вершин. Для птахів, що використають
стійкі струми повітря, характерні більш
широкі, закруглені крила з розбіжними
на кінцях вершинами першорядних махових.
Такий політ використають хижі птахи,
лелеки, пелікани. Широкими колами птахи
поступово набирають висоту й
потім
кружляють, виглядаючи здобич, або,
плануючи із втратою висоти, переміщаються
в потрібному напрямку (мал. 46). Динамічне
ширяння властиве морським птахам
(альбатроси, буревісники, чайки), що
мають довгі, але вузькі, із загостреною
вершиною крила. Використовуючи завихрення
повітря над хвилями або різною швидкістю
повітряних потоків, птах по вітрі планує
вниз, набираючи швидкість, і біля води,
де швидкість вітру вповільнена тертям
об воду, повертається проти вітру й
злітає вгору, де повітря рухається
швидше (мал. 47). Так птах може ширяти
годинниками, виглядаючи здобич і
добуваючи її з пікірування. При
відсутності вітру ці птахи ширяти не
можуть.
Л ітаючі
планіруючим польотом птахи здатні й
до махового польоту. До нього вони
прибігають, щоб знайти висхідний
термічний потік, підлетіти до гнізда,
увернутися від небезпеки й т.п. Однак
довго літати маховим польотом вони не
можуть. З іншого боку, птахи, що літають
маховим польотом, часом переходять на
ковзання або планування. Загалом, кожен
вид користується характерним для нього
польотом, але при необхідності здатний
міняти як характер польоту, так і його
швидкість. Лісові дрібні горобині птахи
літають зі швидкістю 25—40 км/год, голуби
— 30—60, качки й багато куликів — 65—80
км/год; ластівки, що ловлять у повітрі
комах звичайно літають зі швидкістю
40-60 км/год, а стрижі - навіть до 100- 120
км/год. Найшвидший літун нашої фауни
живе у Примор'ї — колючехвостый стриж
— Hirundapus
caudatus
—
розвиває швидкість до170 км у годину.
Наприклад, поштовий голуб, летить зі
швидкістю 50- 60 км/год, здатний пролетіти
без зупинок близько 600 км. На короткий
час птахи здатні розвивати й більші
швидкості. Великі соколи літають зі
швидкістю 60-70 км/ч, але при кидку за
здобиччю розвивають швидкість більше
100 км/ч, а кидаючись на неї з висоти, за
рахунок інерції пікірування - і до
300-350 км/ч. Таку ж швидкість, але на дуже
короткий час - на десятки секунд - здатні
розвивати стрижі, голуби, качки, що
увертаються від хижака.
ітаючі
планіруючим польотом птахи здатні й
до махового польоту. До нього вони
прибігають, щоб знайти висхідний
термічний потік, підлетіти до гнізда,
увернутися від небезпеки й т.п. Однак
довго літати маховим польотом вони не
можуть. З іншого боку, птахи, що літають
маховим польотом, часом переходять на
ковзання або планування. Загалом, кожен
вид користується характерним для нього
польотом, але при необхідності здатний
міняти як характер польоту, так і його
швидкість. Лісові дрібні горобині птахи
літають зі швидкістю 25—40 км/год, голуби
— 30—60, качки й багато куликів — 65—80
км/год; ластівки, що ловлять у повітрі
комах звичайно літають зі швидкістю
40-60 км/год, а стрижі - навіть до 100- 120
км/год. Найшвидший літун нашої фауни
живе у Примор'ї — колючехвостый стриж
— Hirundapus
caudatus
—
розвиває швидкість до170 км у годину.
Наприклад, поштовий голуб, летить зі
швидкістю 50- 60 км/год, здатний пролетіти
без зупинок близько 600 км. На короткий
час птахи здатні розвивати й більші
швидкості. Великі соколи літають зі
швидкістю 60-70 км/ч, але при кидку за
здобиччю розвивають швидкість більше
100 км/ч, а кидаючись на неї з висоти, за
рахунок інерції пікірування - і до
300-350 км/ч. Таку ж швидкість, але на дуже
короткий час - на десятки секунд - здатні
розвивати стрижі, голуби, качки, що
увертаються від хижака.
Можливості й характер польоту кожного виду тісно зв'язані з усіма особливостями його життя: переважним місцеперебуванням, набором кормів і способами його добування, прийомами уникнення небезпеки, дальністю сезонних переміщень і т.п. Види, що годуються у воді й при небезпеці ховаються в заростях, мають швидкий, але не маневрений політ (качки, гагари й ін.). Курячі птахи, що годуючі на землі й при небезпеці намагаються спочатку затаїтися, мають короткі й широкі крила, що дозволяють їм із шумом стрімко злітати під носом приголомшеного хижака й, пролетівши невелику відстань, зникнути в заростях; голосний ляскіт крил при зльоті попереджає про небезпеку сусідів. Швидкий політ рябків дозволяє їм гніздитися далі від водойм й щодня літати на водопій за десятки кілометрів. Розшукуючи здобич, орли, канюки, шуліки годиннами ширяють у повітрі над відкритими просторами. Чатуючі здобич і переслідуючи її в лісі яструби мають маневрений політ, що дозволяє переслідувати здобич у кронах дерев. Розшукуючи здобич на землі й у поверхневих шарах води чайки й крачки чергують махаючий маневрений політ з динамічним ширянням і короткочасним «зависанням». Годинами носяться в повітрі, ловлячи дрібних комах, стрижі; вони чергують швидкий маховий політ з ковзанням. Ці швидкі й невтомні літуни п'ють на льоту, спаровуються в повітрі й навіть здатні вночі спати, повільно, зі швидкістю 30-40 км/год, літаючи колами на великій висоті.
Органи травлення й харчування. Теплокровність і висока рухливість птахів забезпечуються споживанням значної кількості їжі. Але висока рухливість у свою чергу розширює можливості видобутку корму. Загальний спектр кормів, використовуваних класом птахи у цілому, значно різноманітніший, ніж у сучасних плазунів. Більшість птахів тваринноядні. Розмір здобичі визначається величиною птаха й способами її полювання. Великі хижаки здатні успішно справлятися зі здобиччю, що навіть перевищує їх по масі. Багато видів харчуються різноманітними безхребетними, паралельно використовуючи рослинні корми, особливо ягоди й насіння. Гусеподібні, фламінго, що володіють фільтрувальним апаратом дзьоба, харчуються дрібними планктонними тваринами й водоростями. Рослинноядних видів, що харчуються переважно вегетативними частинами рослин, небагато (гусаки, деякі курячі й пастушкові й ін.); всі вони використають і тваринні корми. Багато й всеядних видів. Основний орган захоплення їжі — дзьоб; лише хижі птахи й сови велику здобич схоплюють лапами, а дзьобом її вбивають і розривають. Розмаїтість форм дзьоба в птахів відбиває характер їхньої харчової спеціалізації. Гострий гачок на кінці (хижаки, сови, баклани й ін.) або гострі ріжучі краї дзьоба (чаплі, журавлі) допомагають утримувати велику рухливу здобич. Тонкий пінцетоподібний дзьоб дозволяє витягати дрібних безхребетних з їхніх домівок (комахоїдні горобині) або із ґрунту, зондуючи його (кулики, удод). У горобиних птахів, що харчуються переважно насінням, дзьоб міцний, з ріжучими краями, що дозволяє розгризати оболонки насінь. Потужність дзьоба, забезпечуєьться його кістяковою основою й жувальною мускулатурою, може бути дуже великий. Так, дубоніс — С. соссоthraustes масою всього 50—60 м розкушує кісточки вишень.
У більшості птахів конічний язик займає майже все дно ротової порожнини; у його основній частині розвинені шипики, що допомагають утримувати здобич. У птахів з фільтрувальним апаратом м'ясистий язик забезпечує виштовхування води й ґрунту з ротової порожнини й допомагає проштовхувати в глотку відціджену їжу. Язик насінонядних птахів утримує насіннячко на краю дзьоба при розкусувані. У птахів, що харчуються квітковим соком, (колібрі й ін. ) язик сгортається в трубку, через яку засмоктується нектар. У дятлів язик, що сильно висувається (див. мал. 45), несе на кінці крючковидні шипики, що полегшують витаскування комах з їх вузьких і глибоких ходів. У видів, що ковтають здобич цілком або великими шматками, язик зазвичай малий.
Під язиком у деяких птахів є поглиблення, що може сильно розтягуватися й служити місцем тимчасового зберігання їжі (воронові, в’юрки, пелікани й ін.). У ротову порожнину відкриваються протоки слинних залоз. Слина, змочуючи їжу, полегшує її заковтування. У частини видів липка слина, що покриває язик, полегшує захоплення дрібної здобичі (дятли й ін.). У багатьох ластівок і стрижів саланганов липка слина, що густіє на повітрі, використається й при будівлі гнізд. У деяких птахів у слині присутній фермент амілаза, тому часткове переварювання вуглеводів починається вже з моменту заковтування їжі. За язиком лежить гортанна щілина (веде в трахею), за якої ротова порожнина непомітно переходить у довгу, легко розтяжну трубку - стравохід (мал. 48), що лежить під шкірою шиї. У деяких птахів (курячі, голуби, хижаки, папуги й ін.) у нижній частині стравоходу утворюється об'ємисте розширення - зоб, що служить тимчасовим вмістищем їжі (поки нею переповнений шлунок). У голубів у період розмноження посилено діляться й піддаються жировому переродженню епітеліальні клітини, що відриваються в порожнину зоба й разом з лімфою утворюють так зване молочко (містить білше 10% білка й до 12-15% жиру), яким дорослі птахи вигодовують пташенят.
Шлунок птахів розділяється на два відділи: залозистий (ргоventriсulus) і мускульний (ventriсulus) шлунки. Залозистий шлунок чітко не відмежований від стравоходу, що впадає в нього, і відрізняється лише більш товстими стінками й достатком залоз, що виділяють травні ферменти. Мускульний шлунок має товсті мускульні стінки, а його епітеліальні залози виділяють секрет, що утворює міцну рогоподобну вистилку мускульного шлунка — кутикулу. У видів, що харчуються грубою рослинною їжею, стінки мускульного шлунка особливо тверді, а на товстій кутикулі утворюються бугри. Рясно змочена травними ферментами їжа перетирається в мускульному шлунку завдяки ритмічним скороченням його стінок (до 30 в 1 с). Перетиранню їжі допомагають камінчики, що заковтують птахами, (гастроліти) , які накопичуються в порожнині шлунка й відіграють роль млина. Їх запас регулярно відновляється. У великих рослиноядних видів у мускульному шлунку створюється тиск до 20—30 кг/см2. Таким чином, мускульний шлунок птахів виконує ту ж функцію, що й зуби ссавців при пережовуванні їжі. Дрібно перетерта їжа надходить у кишечник, а неперетравлювані й не здрібнені залишки (волосся, пір'я, кості, хітин) у багатьох птахів збиваються в щільну грудку - погадку - і через стравохід і ротову порожнину викидаються назовні, не захаращуючи кишечник.

Рис. 48. Травна система голуба (по Шимкевичу):
1 — стравохід, 2 — зоб, 3 — залозистий шлунок, 4 — мускульний шлунок, 5 — дванадцятипала кишка, 6 — печінка, 7 — жовчні протоки, 8 — підшлункова залоза, 9 — протоки підшлункової залози, 10 — тонка кишка, 11 — сліпі кншки, 12 — пряма кишка, 13 — клоака, 14 — кишкові брижі (зображене лише її частина), /5 — нирки, 16 — селезінка
Від мускульного шлунка відходить дванадцятипала кишка, вузькою петлею охоплює щільну підшлункову залозу. Двухлопатева печінка має жовчний міхур (немає в ряду видів, у тому числі й у голубів) - резервуар, у якому накопичується жовч. Жовчні протоки, як і протоки підшлункової залози, впадають у дванадцятипалу кишку, що непомітно переходить у тонку кишку. Тонка кишка утворює кілька петель і переходить у коротку пряму кишку, що впадає в клоаку. На границі тонкої й прямої кишок розташовані парні вирости - сліпі кишки, у більшості птахів воони маленьких розмірів. У пташенят розвинений товстостінний сліпий виріст спинної сторони клоаки - фабрицієа сумка, у якій формуються білі кров'яні елементи; у дорослих птахів вона редукується.
Кишечник довше у рослиноядних видів і перевищує довжину тіла в 10 і більше раз (в африканського страуса — в 20 раз); у частини з них добре розвинені й сліпі кишки, у яких їжа піддається переварюванню під впливом власних ферментів і при участі специфічної мікрофлори. У комахоїдних видів кишечник відносно короткий і перевищує довжину тіла лише в 4—6 разів. Інтенсивність травлення в птахів дуже висока. Наприклад, у горобців від заковтування гусениці до виведення з калом неперетравлених залишків проходить лише 15—20 хв, жуків — близько 1год і зерна — 3—4 год. Висока швидкість переварювання забезпечується інтенсивним перетиранням їжі в мускульному шлунку, високою активністю травних ферментів, що працюють при постійній (і високій) температурі тіла, великою поверхнею кишечнику (що зв'язано не тільки з подовженням кишки, але й дуже сильним розвитком ворсинок її слизової оболонки), участю бактеріальної флори.
Високий рівень обміну речовин, що забезпечується інтенсивним травленням, пов'язаний з переробкою великої кількості їжі. Великі тепловтрати у тварин з меншими розмірами тіла в порівнянні з більшими приводять до того, що дрібні види мають потребу у відносно більших кількостях їжі. Маса їжі, що поїдає дрібний горобець за добу, становить 50-80% їхні маси, у більших видів масою 75-100 г (шпаки, дрозди й ін.) - 15-40% і т.п. Добова норма корму залежить і від калорійності їжі, і від температури середовища; похолодання підвищує потребу в їжі. При достатку їжі птахи споживають корму більше, ніж потрібно для покриття поточних витрат, накопичуючи енергетичні резерви переважно у вигляді жиру. Наприклад, яструб тетеревятник, що має масу близько 1,5 кг, задовольняється 150-200 г м'яса на добу, але, піймавши велику здобич, може з'їсти до 800-1000 г. При достатку сарани рожеві шпаки масою 70-80 г можуть з'їсти за добу до 200 г сарани, хоча для підтримки життя їм досить 15-20 г. Здатність до голодування залежить від розмірів птаха і її фізіологічного стану (наявності енергетичних резервів). Дрібні птахи гинуть без їжі вже через 15-30 год, голуби - через 7-9 днів, великі орли й сови можуть голодувати до місяця.
У межах своєї харчової спеціалізації птахи харчуються переважно більш масовими й доступними в даний період кормами. Із цим зв'язана більш-менш чітко виражена у всіх видів птахів сезонна зміна кормів. Особливо різко вона виражена в осілих видів. Наприклад, великий строкатий дятел улітку харчується переважно відкрито живучими комахами, восени добуває їх під корою й у деревині, а взимку харчується насіннями хвойних дерев, роздовбуючи шишки. Більша рухливість і високий рівень вищої нервової діяльності дозволяють птахам швидко виявляти місця концентрації їжі. З масовою появою якогось корму їм можуть харчуватися й птахи з іншим характером харчування. Так, з появою в степу великих кулиг пішої сарани сюди прилітають годуватися качки, кулики, чайки, чаплі, дрібні горобині. Здатність виявляти скупчення їжі й концентруватися на них поряд з інтенсивним травленням і споживанням більших кількостей корму обумовлюють велике значення птахів у біоценозах й їхня роль в обмеженні чисельності багатьох шкідників (комах, гризунів).
О рганы
дихання й газообмін.
Дихальна система птахів різко
відрізняється від дихальних систем
інших наземних хребетних рядом
особливостей, що інтенсифікують подих
і
тим
самим забезпечують високий рівень
споживання кисню.
Через
парні ніздрі повітря засмоктується в
носову порожнину й через хоани переходить
у ротову порожнину. Сюди вузькою щілиною
відкривається гортань, підтримувана
трьома гортанними хрящами. На
відміну
від ссавців верхня гортань птахів не
має голосових зв’язок і не служить
джерелом звуків. За гортанню йде трахея
- гнучка трубка, просвіт якої підтримують
розташовані в її стінках хрящові
трахейні кільця. У порожнині тіла трахея
розпадається на два бронхи, кожний з
яких входить у відповідну легеню й там
розгалужується.
рганы
дихання й газообмін.
Дихальна система птахів різко
відрізняється від дихальних систем
інших наземних хребетних рядом
особливостей, що інтенсифікують подих
і
тим
самим забезпечують високий рівень
споживання кисню.
Через
парні ніздрі повітря засмоктується в
носову порожнину й через хоани переходить
у ротову порожнину. Сюди вузькою щілиною
відкривається гортань, підтримувана
трьома гортанними хрящами. На
відміну
від ссавців верхня гортань птахів не
має голосових зв’язок і не служить
джерелом звуків. За гортанню йде трахея
- гнучка трубка, просвіт якої підтримують
розташовані в її стінках хрящові
трахейні кільця. У порожнині тіла трахея
розпадається на два бронхи, кожний з
яких входить у відповідну легеню й там
розгалужується.
Н ижня
частина трахеї й початкові ділянки
бронхів формують характерну тільки
для птахів нижню гортань (syrinx) - голосовий
апарат, деталі будови якого сильно
варіюють. Джерелом звуків служать
вібруючі при проходженні повітря
перетинки, розташовані між останніми
кільцями трахеї й півкільцями бронхів.
Спеціальна мускулатура міняє натяг
голосових перетинок, змінюючи характер
звуків. Нерідко нижні кільця трахеї
розростаються, утворюючи тонкостінний
кістковий барабан, що підсилює звуки
й міняє їхню модуляцію. В інших видів
резонатором може служити подовжена
трахея, що утворює петлі, що лежать під
шкірою в області зоба або навіть грудини.
ижня
частина трахеї й початкові ділянки
бронхів формують характерну тільки
для птахів нижню гортань (syrinx) - голосовий
апарат, деталі будови якого сильно
варіюють. Джерелом звуків служать
вібруючі при проходженні повітря
перетинки, розташовані між останніми
кільцями трахеї й півкільцями бронхів.
Спеціальна мускулатура міняє натяг
голосових перетинок, змінюючи характер
звуків. Нерідко нижні кільця трахеї
розростаються, утворюючи тонкостінний
кістковий барабан, що підсилює звуки
й міняє їхню модуляцію. В інших видів
резонатором може служити подовжена
трахея, що утворює петлі, що лежать під
шкірою в області зоба або навіть грудини.
Парні легені відносно невеликі по розмірах, досить щільні й мало розтяжні; вони приростають до ребер з боків хребетного стовпа. Увійшовши в легені, бронх розпадається на 15—20 вторинних бронхів (мал. 49, 2), більшість яких закінчується сліпо, а частина з’єднується з повітряними мішками. Вторинні бронхи з’єднуються один з одним численними більш дрібними парабронхами, від яких відходить безліч бронхіол — радіально розташованих коміркових виростів, густо обплетених легеневими кровоносними капілярами. Саме тут іде насичення крові киснем. Загальна дихальна поверхня легенів птахів значно перевищує дихальну поверхню легенів плазунів і цілком порівнянна з дихальною поверхнею легких ссавців. З легенями птахів зв'язані повітряні мішки — прозорі еластичні тонкостінні вирости слизової оболонки вторинних бронхів. Обсяг повітряних мішків приблизно в 10 разів перевищує обсяг легенів. Один з повітряних мішків — межключичный — непарний, чотири парних — шийні, передньо- і задньогрудні, черевні. Повітряні мішки розташовані між внутрішніми органами,а їхні відростки проникають під шкіру й у порожнину великих кісток (плече,стегно й ін.) (мал. 50).
Акт
подиху здійснюється завдяки розширенню
й звуженню грудної клітини. При вдоху,
коли грудина відсувається від хребетного
стовпа, збільшується обсяг порожнини
тіла й еластичні повітряні мішкі
розширюються, засмоктуючи повітря. При
цьому повітря з легенів висасується в
передні повітряні мішки, а повітря із
зовнішнього середовища по трахеї,
бронхам й їхнім розгалуженням іде в
легені й у задні повітряні мішки —
задньогрудні й черевні (мал. 50, 51). При
видиху грудина зміщується до хребетного
стовпа, обсяг порожнини тіла зменшується
й під тиском внутрішніх органів повітря
видавлюється з повітряних мішків.
Утримуючи багато кисню повітря із
задніх повітряних мішків нагнітається
в легені, а повітря з передніх мішків
— міжключичного, шийних і передньогрудних,
що містить уже мало кисню, але багато
вуглекислого газу, проштовхується в
трахею й виводиться назовні. Таким
чином, насичене киснем повітря практично
безупинно, і при вдиху й при видиху,
проходить через легені, збагачуючи
кров киснем (так званий подвійне
дихання). Більш повному насиченню крові
киснем сприяє й рух крові в легенях
назустріч токовища повітря. При
інтенсивному русі, особливо в польоті,
частота дихальних рухів з більшується.
Так, у качки крижня в спокої відбувається
10—16 подихів за 1 хв, а при зльоті 90—120.
У дрібних птахів подих прискорений -
до 60-100 подихів за 1 хв у спокої.
більшується.
Так, у качки крижня в спокої відбувається
10—16 подихів за 1 хв, а при зльоті 90—120.
У дрібних птахів подих прискорений -
до 60-100 подихів за 1 хв у спокої.
Крім інтенсифікації дихання повітряні мішки запобігають перегріву організму при інтенсивному русі, тому що надлишок тепла віддаляється постійно з повітрям. Підвищення внутричеревного тиску при видиху сприяє дефекації. Пірнаючі птахи, збільшуючи тиск у повітряних мішках, можуть зменшувати обсяг і тим самим збільшувати щільність, що полегшує занурення у воду.
Кровоносна система й кровообіг. На відміну від плазунів у птахів повністю роз'єднані велике й мале кола кровообігу: венозні й артеріальні потоки крові ніде не змішуються, права (венозна) половина серця повністю відділена від лівої (артеріальної). Серце четирьохкамерне, із двох передсердь і двох жолудочків. Венозна кров по великих венах збирається в праве передсердя (мал. 52) і переходить у правий жолудочок. Від нього відходить легенева артерія, що розділяється на праву й ліву гілку, по яких венозна кров попадає у відповідну легеню. Артеріальна кров, що окислилася в легенях, по правій і лівій легеневих венах надходить у ліве передсердя. Це мале коло кровообігу.
Велике коло кровообігу починається лівим жолудочком, від якого відходить тільки одна судина — права дуга аорти. Відразу ж після виходу із серця права дуга аорти відокремлює дві судини — праву і ліву безіменні артерії, а сама, круто повернувши над правим бронхом, іде назад уздовж хребетного стовпа як спинна аорта. Кожна з безіменних артерій розділяється на сонну артерію, що йде в голову, і підключичну артерію, що майже відразу ж знову розділяється на плечову артерію, що йде в м'язи крила, і галузисту в м'язах грудини велику грудну артерію. З великих стовбурів, що відходять від спинної аорти, потрібно згадати непарні внутриносну і брижову артерії, що постачають кров'ю шлунок і кишечник, парні стегнові й сідничні артерії, що постачають кров'ю задні кінцівки, м'язи черевної стінки й органи тазової порожнини.
Венозна система птахів схожа на венозну систему плазунів, відрізняючись лише частковою редукцією воротной системи нирок й редукцією черевної вени, функціонально заміщеної куприково-брижовою веною (див. нижче). З голови венозна кров збирається в парні яремні вени, із крила - у плечову вену, із грудних м'язів - у грудну вену. Ці три вени, разом з декількома більш дрібними судинами, з кожної сторони зливаються в короткі й широкі ліві й праві передні порожні вени, що впадають у праве передсердя. Трохи дрібних вен, що збирають кров із клоакальної області, зливаються разом й утворюють три вени: непарну куприково-брижову вену, що проходить по брижам під кишечником і впадає у воротну вену печінки,і парні воротні вени нирок, кожна з яких входить у відповідну нирку. На відміну від плазунів у птахів тільки частина крові розходиться по нирковидних капілярах, а інша кров рухається по великих судинах - продовженням воротних вен нирок - загальним клубовим венам, у які впадають сідничні й стегнові вени. Прийнявши ниркові вени, загальні клубові вени виходять із нирок і зливаються в задню порожню вену (мал. 52), що проходить через печінку, приймаючи в себе печіночні вени, і впадає у праве передсердя.
/ — праве передсердя, 2
—
правий жолудочок, 3
—
легенева артерія, 4
—
легеневі вени, 5 — ліве передсердя, 6
— лівий
жолудочок, 7 — права дуга аорти, 8
—
спинна аорта, 9
—
безіменна артерія, 10
—
загальна сонна артерія, // — підключична
артерія, 12
—
плечова артерія, 13
—
грудна артерія, 14
—
внутриносна артерія, /5 — брижоова
артерія, 16
— стегнова
артерія, 17
—
сіднична артерія, 19
—
яремна вена, 20
—
плечова вена, 21
—грудна
вена, 22 — передня порожня вена, 23
— куприково-брижова
вена, 24
—
воротна вена нирки, 25 — загальна клубова
вена, 26
—
сіднична вена, 27 — стегнова вена. 28
—
ниркова вена, 29
—
задня порожня вена, 30
—
печінкова вена, 31
—
воротна вена печінки, 32
— підшлункова
вена, 33
—
брижова вена
— праве передсердя, 2
—
правий жолудочок, 3
—
легенева артерія, 4
—
легеневі вени, 5 — ліве передсердя, 6
— лівий
жолудочок, 7 — права дуга аорти, 8
—
спинна аорта, 9
—
безіменна артерія, 10
—
загальна сонна артерія, // — підключична
артерія, 12
—
плечова артерія, 13
—
грудна артерія, 14
—
внутриносна артерія, /5 — брижоова
артерія, 16
— стегнова
артерія, 17
—
сіднична артерія, 19
—
яремна вена, 20
—
плечова вена, 21
—грудна
вена, 22 — передня порожня вена, 23
— куприково-брижова
вена, 24
—
воротна вена нирки, 25 — загальна клубова
вена, 26
—
сіднична вена, 27 — стегнова вена. 28
—
ниркова вена, 29
—
задня порожня вена, 30
—
печінкова вена, 31
—
воротна вена печінки, 32
— підшлункова
вена, 33
—
брижова вена
Воротна вена печінки утворюється злиттям згадуваної куприково-брижової вени з декількома венами, що несуть кров від шлунка й кишечника; у печінці вона розпадається на капіляри, кров з яких по печіночних венах виливається в задню порожню вену.
Характерна риса птахів - відносно велики розміри серця; у багатьох маса серця становить близько 1% від маси тіла, а у видів зі швидким маневреним польотом навіть до 1,5-2%; у дрібних видів відносні розміри серця більше, ніж у великих. Висока інтенсивність роботи серця: у птахів середнього розміру (маса близько 0,5 кг) у спокої пульс 200—300 ударів за 1 хв, а в польоті зростає до 400—500; у дрібних птахів у спокої пульс дорівнює 400—600 ударам за 1 хв, збільшуючись у польоті до 1000 і більше. Високий і кров'яний тиск; у птахів різних видів він становить 120—200 мм рт. ст. (у ссавців 70—160 мм, а в плазунів — тільки 30—50 мм). Загальна кількість крові, число еритроцитів і вміст у ній гемоглобіну в птахів помітно вище, ніж у плазунів, і цілком порівнянно із цими показниками в ссавців. Киснева ємність крові в птахів практично така ж, як у ссавців, і вище кисневої ємності крові плазунів в 2-4 рази.
Всі особливості кровоносної системи птахів відповідають високому рівню їхнього метаболізму. Великий обсяг серця й частий пульс створюють швидку циркуляцію крові по організму, що поряд з особливостями крові (висока киснева ємність і значна буферність, велика кількість цукрів), забезпечує безперервне й інтенсивне насичення всіх органів і тканин киснем і живильними речовинами й видалення з них продуктів метаболізму.
Органи виділення й водно-сольовий обмін. Виділення продуктів розпаду й регуляція водного обміну здійснюються переважно нирками. Великі (до 1-2% від маси тіла) метанефричні нирки лежать у поглибленнях тазового пояса. Від кожної нирки відходить сечовід (мал. 53), що відкривається в клоаку. Сечового міхура в птахів немає.
У птахів, як й у більшості плазунів, у якості виділюваного з організму кінцевого продукту білкового обміну утворюється не сечовина, а сечова кислота, що легко випадає з розчину кристаликами, що утворюють білу кашкоподібну масу. Така консистенція виділюваної сечі, обумовила редукцію сечового міхура в птахів. Розміри ниркових клубочків у нирках птахів відносно невеликі, що знижує інтенсивність фільтрації й зменшує втрати води. Основна маса сечової кислоти попадає в просвіт ниркових канальцев не стільки шляхом фільтрації із клубочков, скільки шляхом секреції спеціальними залозистими клітинами стінок ниркових канальцев, що зв’язані капілярами воротної системи нирок. Одночасно ниркові канальці (нефрони) виконують й осморегуляторну функцію. У нефроні птахів з'являється відсутній в анамній й у плазунів 11-образновигнутий середній відділ — петля Генле, густо зв’язана капілярами й функціонуюча як своєрідна «помножуюча противоточня система», що використає осмотичний градієнт — різний зміст солей у тканинах нирки й у просвіті ниркових канальцев. Проникність стінок петлі Генле створює можливість пасивної реабсорбції води з первинної сечі в зонах підвищеної солоності, а іони натрію активно реабсорбуються у висхідній частині петлі й у кінцевій частині канальцев. Цей механізм забезпечує утворення концентрованої сечі. Додатково всмоктування води проходить у клоаці. Все це дозволяє видаляти з організму продукти розпаду при мінімальній втраті води.

А "
Рис. 53. Сечостатеві органи голуба (схема). А -самець; Б -самка:
/ — нирка, 2 — сечовід, 3 — порожнина клоаки, 4 — наднирник, 5 — насінник, 6 — придаток насінника, 7 — сім'япровід, 8 — насінний пухирець, 9 — яєчник, 10 — лівий яйцепровід, //— лійка яйцепроводу, 12 — залишок скороченого правого яйцепроводу, 13 — пряма кишка, 14 — сечовий отвір, 15 — статевий отвір
У більшості птахів є носові залози, розташовані на лобових костях над орбітою. Особливо сильно вони розвинені в морських птахів (трубконосі й ін.) і в деяких пустельних птахів, які змушені пити солону воду. У секреті носових залоз концентрація повареної солі в 4-5 разів вище, ніж у крові, і приблизно вдвічі вище, ніж у морській воді. Виділення настільки концентрованого розчину дозволяє птахам пити воду солоних водойм. Надлишок інших солей виділяється через нирки із сечею.
Статева система й особливості розмноження. Птахи, як і плазуни, - яйцекладущі тварини, але з особливо розвинутою турботою про нащадків. Серед птахів зовсім немає яйцеживородних і живородних видів, що, імовірно, не можна пояснити тільки пристосованістю до польоту.
Парні бобовидні насінники підвішені на брижах біля переднього краю нирок. До початку розмноження в порівнянні з періодом спокою їхній обсяг зростає в 300-1000 разів. До внутрішнього краю кожного насінника прилягає придаток насінника (мал. 53) - залишок мезонефричної нирки. Від кожного придатка насінника починається тонкий сім'япровід (гомологічний вольфову каналу), що проходить по черевній поверхні нирки й впадає в клоаку, утворивши перед цим невелике розширення - насінний пухирець - резервуар, де накопичуються зрілі сперматозоїди.
К опулятивні
органи у вигляді непарної ділянки стінки
клоаки, що вивертається, є лише в деяких
птахів - страусів, тинаму, гусеподібних.
В інших птахів запліднення відбувається
при притисканні зовнішнього отвору
клоаки самця до клоаки самки.
опулятивні
органи у вигляді непарної ділянки стінки
клоаки, що вивертається, є лише в деяких
птахів - страусів, тинаму, гусеподібних.
В інших птахів запліднення відбувається
при притисканні зовнішнього отвору
клоаки самця до клоаки самки.
У самок розвивається тільки лівий яєчник (мал. 53); редукція правого яєчника (і правого яйцепроводу), імовірно, визначається тим, що при великих розмірах' яєць одночасне їхнє формування в парних яєчниках неможливо. Яєчник підвішений на брижах біля переднього кінця лівої нирки. Лівий яйцепровід (мюллєров канал) лійкою відкривається в порожнину тіла біля яєчника, а його стовщений задній (матковий) відділ впадає в ліву частину клоаки. У частини самок зустрічається невеликий сліпий виріст клоаки - залишок скороченого правого яйцепроводу. До початку розмноження частина фолікулів -збільшується в розмірах, тому що ооциты в них інтенсивно накопичують жовток. Одночасно відбувається подовження яйцепроводу й набрякання його стінок. Дозріла яйцеклітина через розрив стінки фолікула випадає в порожнину тіла й, підхоплена сильно збільшеною лійкою, попадає в яйцепровід. Запліднення повинне відбутися в початковій частині яйцепроводу, пов'язаною з лійкою. Після цього яйцеклітина скороченнями стінок яйцепроводу переміщається в напрямку до клоаки, покриваючись декількома оболонками-виділеннями залоз стінок яйцепроводу. Від моменту проникнення яйцеклітини в яйцепровід до повністю сформованого й готового до откладання яйця в різних птахів проходить 12-48 ч.
У вже відкладеному яйці на анімальному полюсі жовтка плаває зародковий диск (мал. 54) - результат дроблення заплідненого ядра яйцеклітини й навколишнього його тонкого шару протоплазми. Жовток оточений тонкою жовтковою оболонкою, що сформувалася з виділень залоз стінок початкової частини яйцепроводу. Далі йде товста білкова оболонка, що складається з декількох шарів - більш густих і більш рідких. Зовні білкова оболонка покрита двома тонкими пергаментоподібними підшкорлуповими оболонками, які на тупому кінці яйця розходяться, утворюючи повітряну камеру. Від внутрішньої підшкорлупової оболонки до жовтка йдуть звиті джгути щільного білка - халази; вільно повертається на скручуючихся халазах жовток при будь-якому положенні яйця має зародковий диск нагорі. Зовнішня оболонка яйця - щільна вапняна шкарлупа.
Співвідношення жовтка й білка в яйцях різних груп птахів варіює. Так, у горобиних птахів жовток становить 10—25%, а білок 70—80% від маси яйця, у гусаків маси жовтка й білка подібні й становлять приблизно по 44% від маси яйця. Жовток служить основним запасом живильних речовин, які йдуть на формування тканин зародка й забезпечують основні енергетичні витрати, а частково - і потребу у воді, білкова оболонка служить в основному джерелом необхідної для розвитку зародка води й додатковим резервом енергетичних речовин. Хімічний склад білка й жовтка варіює в різних груп. У середньому в яйці птахів утримується (%):
У жовтку В білку
Води 40—56 85—90
Жирів і липоїдів 30—40 1—3
Протеїнів 15—20 10—15
Вуглеводів Сліди 0,5—1
Мінеральних речовин………… 1—3 0,5—1,5
Шкарлупа на 92—95% складається із кристалів вуглекислого кальцію, невеликої кількості вуглекислого магнію, фосфорнокислих кальцію й магнію й 3—5% органічних речовин. У дрібних птахів маса шкарлупи становить 5—10% від загальної маси яйця, у великих і важких птахів шкарлупа товще й може становити до 15—20% маси яйця. Найтонші пори, що пронизують шкарлупу, забезпечують доступ кисню до зародка, що розвивається. У міру розвитку зародка частина солей зі шкарлупи переходить у його кров'яне русло й використовується на формування кістяка. При цьому вміст солей: усередині яйця зростає в 4—5 разів, а шкарлупа стає більш крихкою, що полегшує народження пташеняти. У більшості птахів шкарлупа містить пігменти, які виділяються залозистими клітинами яйцепровіда. Забарвлення може бути однотонним або плямистим. У частини наземних видів шкарлупа забарвлена під тло субстрату. У багатьох видів, що відкладають яйця в дупла й нори, шкарлупа біла.
У самих дрібних птахів — деяких колібрі, що мають масу 1,6—1,8 гр, маса яйця близько 0,2 гр, в африканського страуса й великих пінгвінів маса яйця становить близько 1,5% маси самки, а в куликів і дрібних горобиних птахів — до 15—20%. Плідність птахів помітно менше плідності плазунів, що пов'язане зі зменшенням ембріональної й постембріоналної смертності завдяки різноманітним формам турботи про нащадків. Великі хижаки, пінгвіни, кайри й дрібні види колібрі кладуть одне яйце. У голубів, стрижів, дрібних пінгвінів, великих колібрі, журавлів, дрімлюг у кладці два яйця. Три яйця відкладають рябки й більшість чайок. У куликів у кладці чотири яйця. Більшість горобиних відкладають по 5—8 яєць, качки — по 6—14 яєць і т.п. У денних хижаків і сов розміри кладки зростають при достатку їжі. Самі великі кладки — до 22 яєць — бувають у сірої куріпки й переспівала. Більші кладки — до 10—15 яєць — відомі для деяких дрібних горобиних: синиць, кропив'яників і ін.
Розмноження кожного виду птахів приурочене до певного сезону року так, що ріст пташенят відбувається, у найбільш їстівний період. У помірних і північних широтах розмноження починається наприкінці весни - першій половині літа, у тропіках воно приурочене до періоду дощів або, навпаки, до періодів посухи. У помірних і північних широтах збільшення довжини світлового дня викликає секрецію гіпофізом гонадотропних гормонів, під впливом яких у полових залозах починається формування статевих клітин. Остаточне дозрівання статевих клітин і безпосередній початок розмноження (запліднення й відкладання яєць) визначається комплексом факторів: сприятливими температурами, достатньою кількістю корму, наявністю підходящих місць для гніздування, присутністю статевого партнера.
У всіх птахів у тому або іншому ступені виражений статевий диморфізм. Найбільш чітко він проявляється в багатьох курячих, гусеподібних, горобиних, коли самці помітно крупніше самок, більш яскраво забарвленні, іноді мають більш гучний голос. В інших птахів він виражений лише в тому, що самці трохи крупніше самок (у хижих птахів і сов самки крупніше самців). У більшості видів самець і самка на період розмноження утворюють пари; в орлів, лебедів, гусаків, великих чапель пари зберігаються кілька років, іноді до загибелі одного з партнерів, в інших, наприклад у багатьох качок, лише в період откладки яєць. Всіх цих птахів називають моногамами. У меншого числа видів - полігаморів - пари не утворюються навіть на короткий час і спарювання відбувається при короткочасних зустрічах самців і самок; звичайно в них особливо різко виражений статевий диморфізм. До полігаморів відносяться глухари, тетерева, павичі, кулики турухтани, колібрі й ін.
Розмноження у всіх птахів починається шлюбними іграми або токовищем. Його прояви надзвичайно різноманітні: це й звучна пісня горобиних птахів, і танцю журавлів, і барабанний дріб дятлів, що стукають дзьобом по сухому сучку, і голосні лементи сов, і тяга вальдшнепа й т.д. У моногамів у шлюбних іграх у тім або іншому ступені беруть участь обоє партнера, але самець токует більш інтенсивно. У деяких видів, наприклад у куликів плавунчиків, токуют самки; вони крупніше самців й яскравіше забарвленні. У полігаморів токуют тільки самці. Токові явища полегшують зустріч самця й самки, сприяють формуванню пари (у моногамов), забезпечують фізіологічну підготовку партнерів до спарювання. Чітко виражена видоспецифічність токових явищ (поз і загального характеру поводження, видаваних звуків) запобігає міжвидовій гібридизації.
У період токовища йде й будівля гнізда. У моногамів його будують обоє партнера або тільки самка, а самець іноді підносить будівельний матеріал. У полігаморів гніздо будує тільки самка. Характер гнізд й їхнє розташування в класі птахи надзвичайно різноманітне (мал. 55), але в кожного виду відповідно до його екологічних особливостей гнізда відносно однотипні. Лише далеко не всі птахи гнізд не будують, відкладаючи яйця прямо на землю (дрімлюги, деякі кулики) або на уступи скель (кайри). Устеляють лунку на землі підстилкою з рослинного дрантя курячі й кулики, чайки, сови й ін. Після завершення кладки самки качок і гусаків, вискубуючи в себе на черевці пух, вистилають їм гніздо; при відході на годівлю закривають яйця пухом.

А — кладка на землі без гнізда в дрімлюги й Б — кулика зуйка; В — гніздо на землі рябчика й Г — звичайної чайки; Д — яйце кайри на голій скелі; Е — гніздо мойовки на скелястому уступі; Ж — імператорський пінгвін з яйцем на лабетах; 3 — зліплене із бруду гніздо сільської ластівки; И — вирита щуриками гніздова нора; гнізда на воді: К — великої поганки й Л — чорної крачки; гнізда на деревах: М — горлиці; Н — зяблика; О — славки-кравчині; П — спільне гніздо африканських горобців; Р — великий строкатий дятел у видовбаного гніздового дупла
У розналужені гілок недбалі гнізда з накиданих сухих гілок з убогої трав'янистої вистилки влаштовують денні хижаки, лелеки, чаплі, баклани, воронові; іноді вони будують їх у заломах очерету, на скелястих уступах або на землі. Особливо мистецьки будують гнізда багато горобиних птахів, поміщаючи їх у розгалужені гілок або на землі. Щільна чаша гнізда звивається із сухих травинок і тонких гілочок, а зсередини вистилається м'якими стеблинками, мохами, підібраним пір'ям і вовною. Деякі птахи, наприклад зяблики, для маскування гнізда вплітають у його зовнішні стінки лишайники й шматочки кори. Співочі дрозди промащують лоток змоченою слиною порохнявою деревиною, а інші дрозди зміцнюють стінки брудом. У деяких видів гніздо являє собою щільну кулю з товстими стінками й бічним входом, побудований у розгалужені гілок (кропив'яник, довгохвоста синиця) або підвішений до тонких гілочок дерева (синиця ремез, багато тропічні тка-чики й ін.).
Дятли довбають дупла, на дно яких без усякої підстилки відкладають яйця. Дятлові й природні дупла займають багато птахів, що влаштовують на дні гніздо: деякі качки, голуби, сови, стрижі, різні горобині. Самці птахів носорогів замазують вхід у дупло брудом, залишаючи лише невеликий отвір, через який годує самку, що насиджує. Багато ластівок ліплять гніздо із грудочок бруду, скріплюючи їхньою липкою слиною. Стрижі салангани влаштовують гніздо тільки зі швидко густіючої на повітрі слини. Дрібні трубконосі, тупики й сокирки, зимородки, щурки, щурики й інші птахи риють нори довжиною до 1-2 м і навіть більше; наприкінці нори будується гніздо або яйця відкладаються прямо на землю. У степах і пустелях каменки, огари й пеганки гніздяться в норах гризунів і вимоїнах обривів. Деякі птахи (лисухи, поганки, деякі крачки) влаштовують плаваючі гнізда із сухих стебел і водоростей, розміщаючи їх у заростях надводної рослинності.
Гніздо захищає кладку, птаха, що насиджує, й пташенят, що вилупилися, від ворогів і несприятливих погодних умов. Захисні якості гнізда тим вище, чим краще воно побудовано й чим більш воно недоступніше. Так, у Підмосковських лісах, де буває багато людей і ведеться випас худоби, гине до 40-50% гнізд, розташованих на землі й низьких кущах, до 20-30% гнізд на деревах і лише 5-10% гнізд у дуплах. Часте лякання птахів, що насиджують, підвищує загибель яєць і пташенят, тому що полегшує виявлення гнізд різними хижаками (воронами, сороками, кішками й ін.). Гнізда істотно поліпшують умови інкубації, тому що коливання температури в них значно менше, ніж у зовнішнім середовищі.
Практично всі птахи насиджують яйця, тобто обігрівають їх. Не насиджують кладок лише бур'янисті кури — Megapodiidae: необхідне для розвитку зародка тепло утворюється при гнитті рослинного дрантя об'ємистого «гнізда». У полігаморів насиджують тільки самки; у моногамів у насиджуванні беруть участь обоє партнера, що міняють один одного на гнізді (чайки, багато горобиних й ін.), або насиджує тільки самка, а самець її годує й охороняє район гнізда. У птахів, що насиджують, на черевці випадає пір'я й пух і утворюється насідла пляма - ділянка голої шкіри із сильно розвиненими кровоносними судинами, яким птах притискається до яєць. Насідних плям буває 2—3 або одна велика. Теплоізоляційні якості гнізда й наявність насідних плям забезпечує гарне прогрівання яєць: температура кладки досягає 36—38° С. Не утворюється насідних плям у гусеподібних, але їхня відсутність компенсується щільною пуховою вистилкою гнізда. Імператорські пінгвіни, що гніздяться на льодах Антарктиди, тримають своє єдине яйце на лабетах, прикриваючи його зверху складкою шкіри черева; при температурі повітря в-5-10 С температура усередині яйця дорівнює +36-37 С.
Більшість птахів починає інтенсивне насиджуваня після відкладання всіх яєць, і тому вилуплення пташенят проходить більш-менш одночасно. У денних хижаків, сов і деяких інших птахів насиджування починається після відкладки першого яйця. Відповідно вилуплення пташенят йде поступово й при більших кладках розтягується на 5-10 днів. Тривалість інкубаційного періоду залежить від розмірів яйця й птаха, від типу гнізда й інтенсивності насиджування. Дрібні горобині насиджують 11-14 діб, ворона- 17, ворон - 19-21, чирок трескунок - 21-24, крижень - 26, лебеді 35-40 доби й т.д. Найбільша тривалість насиживания - біля двох місяців - у великих пінгвінів, альбатросів, грифів.
Розвиток. В період відкладки яйця вже утворюється плаваючий на жовтку зародковий диск. Подальший розвиток відновлюється лише після початку насиживания.У центральній частині зародкового диска утворюється стовщення - первинна смужка, у якій відбувається формування трьох зародкових листків: ектодерми, мезодерми й ентодерми. Далі йдуть два паралельних процеси: розростання країв зародкового диска, що приводить до утворення зародкових оболонок і до обростання жовтка, і розвиток зародка, що формується в області первинної смужки.
Уже в першу добу з ентодерми формується хорда, а лежача над нею смужка ектодерми перетворюється в нервову трубку, на передньому кінці якої з'являються здуття - мозкові міхури. З боків хорди й нервової трубки мезодерма диференціюється на соміти й починають формуватися кровоносні судини й серце, травна трубка з п'ятьма парами зябрових щілин в області глотки, які незабаром заростають, і т.д. Зародок відокремлюється від периферичних ділянок зародкового диска. Далі утворюється дзьоб, з'являються зачатки кінцівок. На вторий-третій день інкубації, одночасно з формуванням кровоносних судин у тілі зародка, густа мережа капілярів - судинне поле - утворюється в периферійних частинах зародкового диска. Вони зливаються в жовточні вени,

Рис. 56. Поздовжній розріз курячого яйця на різних стадіях інкубації (І—IV)
(по Заварзіну):
/ — зародок (розріз поперек тіла), 2 — жовток, 3 — білок, 4 — амніотична складка, 5 — позазародкова порожнина, 6 — повітряна камера яйця, 7 — шкарлупа, 8 — серозна оболонка, 9 — амніон, 10 — амніотична порожнина, 11 — аллантоїс, 12 — жовточний
мішок
що впадають в серце; по них живильні речовини жовтка надходять у кровоток зародка. Одночасно формуються зародкові оболонки (мал. 56) - серозна й амніотична. Як виріст задньої кишки зародка утворюється аллантоїс, що вростає між амніоном і серозною оболонкою.
До кінця інкубаційного періоду жовточний мішок із залишками жовтка втягується в черевну порожнину, зародок прориває зародкові оболонки й, висунувши дзьоб у повітряну камеру, починає дихати легенями; кровопостачання аллантоїса припиняється. Потім пташеня за допомогою «яйцевого зуба» — вапняного горбка на кінці дзьоба— надкльовує зсередини шкарлупу, поступово розширює щілина й виповзає зі шкарлупи й зародкових оболонок. Від прокльова шкарлупи до виходу з яйця в дрібних птахів проходить кілька годин, у більших — 1—3 доби. При вилуплені дорослий птах сидить на гнізді, обігріваючи кладку.
По ступені фізіологічної зрілості пташеняти в момент вилуплення всіх птахів можна розділити на дві групи: зріловилупляються - матуронатні (виводкові) - і незріловилупляються - імматуронатні (пташеняткові). Матуронатні пташенята (мал. 57) опушені й

зрячі; обсохнувши, вони можуть добре бігати, а водні — плавати й навіть пірнати; незабаром після вылуплення вони залишають гніздо й кочують, самостійно харчуючись. Роль дорослого птаха зводиться до захисту виводка, періодичному обігріву пташенят (терморегуляція проявляється вже в перші 1-3 дні після вилуплення) і допомоги в пошуках їжі. Наслідування дорослому птахові, що водить виведень, дозволяє пташенятам виробити навички пошуку й схоплювання здообичі, відхилення від небезпеки й т.п. До цієї групи відносяться наземні й водні птахи: страуси, тинаму, гусеподібні, куроподібні (за винятком гоацина), журавлі, пастушки, дрохви, більшість куликів.
Імматуронатні пташенята виходять із яйця безпомічними,, сліпими, голими або слабко опушеними (мал. 57). У гнізді залишаються довго: у горобиних птахів - 10-12 днів, у трубконосих - до 2 місяців і більше; вигодовуються й обігріваються батьками. У перші дні життя імматуронатні пташенята на будь-яке подразнення відповідають однозначно вимогою корму: відкривають дзьоб і пищать. Лише пізніше, коли в них прорізаються очі й відкриваються зовнішні слухові проходи (на 4-7-й день), їх поведінка ускладнюється; вони вимагають корм лише з появою в гнізда дорослих птахів, а при будь-яких інших подразненнях затаюються на дні гнізда. У першу половину гніздового життя імматуронатні пташенята пойкілотермні: їх температура цілком залежить від температури середовища. Тому дорослі птахи інтенсивно обігрівають пташенят. Якщо ж дорослі птахи надовго залишають гніздо, то, особливо при холодній погоді, температура тіла пташеняти знижується, зменшується число подихів і пульс і пташенята впадають у стан анабіозу, не реагуючи на подразнення. Дорослий птах, що повернувся, нагріє пташенят, рівень обміну речовин й їхня активність знову зростуть. Ця особливість забезпечує можливість швидкого росту пташеняти, тому що живильні речовини корму не витрачаються на терморегуляцію, а йдуть тільки на ріст тіла. При інтенсивній годівлі й обігріві за добу пташенята в багатьох видів збільшують масу на 20-50%.
Інтенсивність вигодовування пташенят в більшості видів велика. Так, пари великих строкатих дятлів своїм 5-6 пташенятам за добу приносить 150-250 порцій корму, пари великих синиць приносили своїм 14 пташенятам до 500 порцій корму в день і т.п. У перші дні батьки носять пташенятам м'яку, легко засвоювану їжу (синиці в перший-другий день годують пташенят павуками). У другій половині гніздового життя в пташенят розвертається оперення й поступово встановлюється терморегуляція. Вони залишають гніздо опереними, уже досягши маси й розмірів дорослих птахів, активним, хоча ще невпевненим польотом. Після вильоту батьки якийсь час (1-2 тижня) продовжують годувати пташенят. У цей час, наслідуючи дорослих, пташеняти виробляють властивий даному виду стереотип поведінки: прийоми пошуку й схоплювання здобичі, реакції на небезпеку й т.п. До імматуронатних належать голуби, папуги, стрижеподібні, дятлоподібні, горобині; з водних птахів типово імматуронатними будуть тільки пеліканоподібні.
Досить багато птахів становлять проміжну групу. Пташенята гагар і поганок лупляться опушеними й зрячими, здатними відразу ж добре плавати й пірнати, але батьки їх годують до підйому на крило. Опушені й зрячі пташенята більшості чайок швидко залишають гнізда, але батьки годують їх і після підйому на крило. У трубконосих і більшості чистикових птахів оперені й зрячі пташенята залишаються в гнізді й одержують корм від батьків, поки не досягнуть розмірів дорослих і не надбають здатність до польоту. У сов і денних хижаків пташенята лупляться опушеними і сліпими, але прозрівають швидко, іноді в день вилуплення; вони теж вигодовуються бітьками в гніздах до підйому на крило. У ряді лелекоподібні у чапель пташенята лупляться голими, уже після вилуплення прорізаються очі й розвивається ембріональний пух, після чого пташенята при небезпеці можуть залишати гніздо й ховатися поблизу. У лелек пташенята лупляться зрячими й покритими пухом, у гнізді залишаються до підйому на крило. Таким чином, у проміжній групі представлені всі переходи між матуро-натним й імматуронатним типами розвитку.
Імовірно, первинним для птахів був матуронатний тип розвитку, що відрізняється від розвитку плазунів лише тим, що турбота дорослих про пташенят різко зменшувала постембріональну смертність, а періодичні обігріви й допомога в пошуках їжі сприяли прискоренню росту. Пізніше виникла імматуронатність прискорила постембріональний ріст завдяки інтенсивному вигодовуванню й пізньому надбанню терморегуляції. Тому імматуронатні птахи швидше завершують свій розвиток. Наприклад, пташенята воронових досягають розмірів дорослих і здобувають здатність до польоту у віці 20-30 днів, а в подібних з ними по розмірах матуронатних птахів - лише у віці 40-60 днів (гусеподібні, пастушкові й ін.).
 Гніздовий
паразитизм.
Приблизно в 80 переважно тропічних видів
птахів: близько 50 видів зозуль - Cuculidae
, 12 видів медоуказчикiв
- Indicatoridae,
близько 5 видів трупіалів - Icteridae,
10 видів африканських ткачиків - Ploceidae
й південноамериканської качки
— Heteronetta atricapilla
— виражений
гніздовий паразитизм: вони не будують
гнізд, підкладають свої яйця в гнізда
інших, частіше близьких видів птахів,
надаючи їм і насиджування яєць, і
вигодовування пташенят.
Гніздовий
паразитизм.
Приблизно в 80 переважно тропічних видів
птахів: близько 50 видів зозуль - Cuculidae
, 12 видів медоуказчикiв
- Indicatoridae,
близько 5 видів трупіалів - Icteridae,
10 видів африканських ткачиків - Ploceidae
й південноамериканської качки
— Heteronetta atricapilla
— виражений
гніздовий паразитизм: вони не будують
гнізд, підкладають свої яйця в гнізда
інших, частіше близьких видів птахів,
надаючи їм і насиджування яєць, і
вигодовування пташенят.
У фауні СРСР представниками цієї групи птахів будуть звичайна й глуха зозулі—Cuculus canoras і С. optatus. Яйця зозуль відносно малі (близько 3% маси самки), а забарвлення яєць, що відкладають різними самками, дуже різна. Це дозволяє використовувати для підкладки гнізда великої кількості видів дрібних птахів. Звичайно «прийомні батьки» не звертають уваги на підкладене яйце, особливо якщо воно по забарвленню схоже, на їхні яйця, і продовжують насиджувати кладку. Зозуленя, що вилупилося з яйця, при охолодженні, коли дорослих птахів немає на гнізді, не знижує активність, як пташенята всіх горобиних птахів, а, навпаки, енергійно рухається по гнізду, намагаючись підлізти під пташеня або яйце хазяїв і потім, різко піднімаючись, викидає його із гнізда (мал. 58). Залишившись один у гнізді, зозуленя одержує більше корму, швидко росте, опушується й у віці 20 днів залишає гніздо, майже досягши - маси дорослої зозулі (близько 100 г); ще біля двох тижнів він невідступно слідкує за прийомними батьками, продовжуючи випрошувати в них корм. Інші види птахів — гніздових паразитів частіше мають вузьке коло хазяїв і підкладають яйця лише одного виду. Їхні пташенята звичайно не здатні викидати інших пташенят і виховуються разом із пташенятами прийомних батьків. Відносно невелике число видів типових гніздових паразитів свідчить, що таке своєрідне розмноження не дає яких-небудь особливих переваг у боротьбі за існування. Початкова фаза гніздового паразитизму — заняття чужих гнізд — зустрічається у видів з різних рядів (деякі хижаки, кулики, дятли й ін.).
Тривалість життя. Смертність молодих птахів у перший рік життя досить висока й часто становить 30—50 і більше відсотків від числа піднявшихся на крило. Вона визначається недостатнім індивідуальним досвідом, що не дозволяє молодим настільки ж успішно, як дорослим, знаходити корм й уникати небезпек. У дорослих птахів різних видів смертність становить 5—20—40% у рік. Зараз для багатьох видів визначальним фактором стала господарська діяльність людини. Багато видів — об'єкти полювання — різко знизили чисельність через непомірне переслідування. На чисельності багатьох видів позначається й непрямий вплив людини: зміна ландшафтів, забруднення вод нафтою й промисловими стоками й т.п. Багато птахів гине при масовому застосуванні гербіцидів й інсектицидів. Більшість дрібних і середніх по розмірах птахів стають статевозрілими й приступають до розмноження на наступне літо,

Рис. 59. Головний мозок голуба (по Паркеру): А — зверху з очними яблуками, розкрита кришка правої півкулі; Б — знизу; В — збоку:
/ — великі півкулі, 2 — смугасте тіло правої півкулі, 3 — нюхові частки, 4 — епіфіз 5 — гіпофіз, 6 — хіазми й зорові нерви, 7 — зорові частки середнього мозку, 8 — мозочок, 9 — довгастий мозок, 10 — очі
у віці 9-10 місяців. Великі воронові (ворон, сіра ворона), дрібні хижаки й чайки вступають у розмноження у віці біля двох років, великі чайки, орли, трубконоси - у віці 3-4 років, страуси - 4-5 років і т.п. Даних про тривалість життя птахів у природних умовах поки мало. Гранична тривалість життя навіть у дрібних горобиних птахів досить висока й досягає 10-20 років, у чайок - 36, у буревісників - 30, а в альбатросів - 40 років. Але середня тривалість життя звичайно менше. Так, на Куршской косі (Калінінградська обл.) половину популяції зябликів, що гніздяться, становлять птаха у віці біля року, а на кожну тисячу птахів, що гніздяться, доводиться тільки три особини старше 7 років. У схемі чим більше тривалість життя, тим менше плідність і тим пізніше наступає статева зрілість. Пряма залежність між розмірами тіла й тривалістю життя в птахів проявляється далеко не завжди: у природі гранична тривалість життя великих лебедів, менше, ніж у більш дрібних чайок і трубконосих. Це визначається видовими особливостями екології, характером впливу людини й ворогів, впливом погодних умов і багатьма іншими факторами.
Нервова система, органи чуття. У будові головного мозку птахів (мал. 59) і плазунів багато загального. В обох класів значну частину добре відособлених великих півкуль переднього мозку становлять смугасті тіла (corpora striata) - розростання дна переднього мозку. У тонкому даху великих півкуль є скупчення нервових клітин, позначуваних як первинна мантія - архіпалірозум - і гомологи елементів неопаліума. Добре розвинений середній мозок, разом зі смугастими тілами забезпечують складні форми нервової діяльності в обох класах. Добре виражений вигин мозку.
Однак у будові головного мозку птахів є й риси більше високої організації. Насамперед помітно зростає загальна маса головного мозку. У плазунів маса головного мозку становить 0,01—0,4% від маси тіла, у безкільових птахів і пінгвінів 0,04— 0,09, а в інших літаючих птахів — від 0,2 до 5—8% від маси тіла. Маса головного й спинного мозку в плазунів приблизно однакова, у птахів маса головного мозку більше (у курячих 1,5:1, у голубів 2,5:1). У плазунів передній мозок становить 42—52% від маси всього головного мозку. У птахів відносні розміри переднього мозку зростають й у більшості рядів становлять 52—62%, а в дрібних горобиних —до 70% маси всього головного мозку (М. Ф. Никитенко, 1969).
Нюхові частки малі й тісно примикають до великих півкуль. На спинній поверхні проміжного мозку, прикритого зверху півкулями, лежить слабко розвитий епіфіз, а на дні - за хіазми зорового нерва - великий гіпофіз. Великі зорові частки середнього мозку відсунуті в сторони півкулями, що налягають на середній мозок, і мозочком. Великий мозочок впритул примикає до великих півкуль, прикриваючи середній і значну частину довгастого мозку. Він має складну складчасту, будова. Мозочок - центр координації рухів і рівноваги - добре розвинений у всіх птахів. Довгастий мозок плавно переходить у спинний. У птахів 12 пар головних нервів (але одинадцята пара ще не чітко відмежована від десятої пари), У спинному мозку птахів розвинені плечові й поперекові стовщення, а спинномозкові нерви, що відходять від них, при виході із хребців утворюють плечові й поперекові сплетення, гілки яких іннервують мускулатуру відповідних кінцівок. Великий головний мозок з більш ускладненою будовою (особливо гістологічним) і відповідні йому органи чуттів забезпечують птахам більш високий у порівнянні із плазунами рівень нервової діяльності й більше складні форми поводження.
Зір
— основний рецептор далекої й ближньої
орієнтації птахів. На відміну від інших
хребетних серед них немає жодного виду
зі скороченими очами. Очі дуже великі
по відносних й абсолютних розмірах: у
великих хижаків і сов у обсязі вони
рівні оку дорослої людини. Збільшення
абсолютних розмірів око вигідно тому,
що дозволяє одержати більші розміри
зображення, на сітківці й тим самим
ясніше розрізнити його деталі. Відносні
розміри очей, що відрізняються в різних
видів, пов'язані з характером харчової
спеціалізації й способами полювання.
У переважно рослиноядних гусаків і
курей очі по масі приблизно дорівнюють
масі головного мозку й становлять
0,4—0,6%
від маси тіла, у тих що ловлять рухливу
здобич і хижих птахів маса ока в 2-3 рази
перевищує масу мозку й становить 0,5-3%
від маси тіла, в активних у сутінках і
вночі сов маса  ока
дорівнює 1—5% маси тіла (М.
Ф.
Никитенко).
ока
дорівнює 1—5% маси тіла (М.
Ф.
Никитенко).
У різних видів на 1 мм2 сітківки знаходиться від 50 тис. до 300 тис. фоторецепторів — паличок і ковбочок, а в області гострого зору — до 500 тис. — 1 млн. При різному сполученні паличок і ковбочек це дозволяє або розрізняти багато деталей об'єкта, або його контури при низькій освітленості. Основний аналіз зорових сприйняттів проводиться в зорових центрах головного мозку; гангліозні клітини сітківки реагують на кілька стимулів: контури, колірні плями, напрямки переміщень і т.д. У птахів, як й інших хребетних, на сітківці є ділянка найбільш гострого зору з поглибленням (ямкою) у його центрі. У деяких видів, що харчуються переважно рухливими об'єктами, є дві області гострого зору: у денних хижаків, чапель, зимородків, ластівок; у стрижів лише одна область гострого зору, і тому їхні способи ловлі здобичі на льоту менш різноманітні, чим у ластівок. У ковбочках перебувають масляні краплі - кольорові (червоні, жовтогарячі, блакитні й ін.) або безбарвні. Імовірно, вони виконують роль світлофільтрів, що підвищують контрастність зображення. Дуже рухлива зіниця запобігає зайвому засвіченю сітківки (при швидких поворотах у польоті й т.п.).
Акомодація (наведення ока на різкість) здійснюється зміною форми хришталика і його одночасним переміщенням, а також деякою зміною кривизни роговиці. В області сліпої плями (місця входження зорового нерва) розташований гребінь — багате судинами складчасте утворення, що вдається в склоподібне тіло (мал. 60, 13). Основна його функція - постачання склоподібного тіла й внутрішніх шарів сітківки киснем і видалення продуктів метаболізму. Гребінь є й в очах плазунів, але в птахів, у зв'язку з більшими розмірами очей, він значно крупніше й складніше. Механічна міцність великих очей птахів забезпечується стовщенням склери й появою в ній кісткових пластинок. Добре розвинені рухливі віка, у деяких птахів несучі вії. Розвинено мигальну перетинку (третє віко), що рухається безпосередньо по поверхні роговиці, очищуючи її.
У більшості птахів очі розташовані з боків голови. Поле зору кожного ока становить 150—170°, але поле бінокулярного зору невелике й становить у багатьох птахів лише 20—30°. У сов й

Рис. 61. Поля зору в яструба (по Уоллсу):
/ — поле бінокулярного зору, 2 — поле монокулярного зору, 3 — область поза полем зреиня, 4 — область проекції обох скроневих ділянок гострого зору, В — проекція центральної ділянки гострого зору
деяких хижих птахів ока зміщаються до дзьоба й поле бінокулярного зору зростає. У деяких видів з опуклими очами й вузькою головою (деякі кулики, качки й ін.) загальне поле зору може бути 360°, при цьому вузькі (5—10°) поля бінокулярного зору утворюються перед дзьобом (полегшує схоплювання здобичі) і в області потилиці. У птахів із двома областями гострого зору вони звичайно розташовані так, що одна з них проектується в область бінокулярного зору, а інша - в область монокулярного зору (мал. 61).
Усі птахи мають кольоровий зір, розпізнаючи не тільки основні кольори, але і їхні відтінки й сполучення. Тому в оперенні птахів так часто зустрічаються яскраві кольорові плями, що виконують функцію видових міток. Гострота зору птахів, перевершує в кілька разів гостроту зору людини. Наприклад, сокіл сапсан бачить здобич, що рухається, розміром з галку на відстані біля 1 км. Птахи розрізняють не тільки переміщення предметів й їхні контури, але й деталі форми й забарвлення, малюнок і фактуру поверхонь. Тому зорові сприйняття використовуються птахами й для одержання різноманітної інформації про навколишній світ, і як важливий засіб при внутрішньовидовому й міжвидовому спілкуванні.
Орган слуху, подібно органу зору, служить у птахів важливим рецептором орієнтації й спілкування. Анатомічно орган слуху подібний з органом слуху плазунів, особливо крокодилів, але завдяки дрібним перетворенням функціонально він не відрізняється від значно більш складного й диференційованого органа слуху ссавців. Внутрішнє вухо птахів відрізняється від внутрішнього вуха крокодилів лише трохи кращим розвитком равлика - подовженого виросту круглого мішечка - і більш складною внутрішньою її будовою. Зростають розміри порожнини середнього вуха, а єдина слуховая кісточка - стремінце - має ускладнену форму, що збільшує її рухливість при коливаннях барабанної перетинки, що має в птахів куполоподібну форму й більші розміри. Барабанна перетинка занурена нижче рівня шкіри й до неї веде канал - зовнішній слуховой прохід, по краю якого в частини видів птахів утворюється складка шкіри - зачаток зовнішнього вуха (добре розвинений у сов). Контурні пір'я, що прикривають зовнішній слуховой прохід, за структурою відрізняються від пір'я прилеглих ділянок голови й служать не тільки для механічного захисту слухового проходу, але й для організації звукового потоку (можуть піднімати, виконуючи роль рупора або, навпаки, притискатися і т.п.).
Птахів з погано розвиненим слухом немає. Більшість видів чує у великому діапазоні — від 30 до 20 тис. Гц, тобто приблизно в діапазоні загостреного людського слуху; деякі види, імовірно, здатні сприймати й ультразвуки до 35—50 кгц, присутні й у їхньому голосі. У цьому діапазоні орган слуху проявляє особливу чутливість до біологічно важливих для даного виду звукам (сигналам свого виду, звукам, видаваним більш звичайними харчовими об'єктами або ворогами, і т.п.). Багато птахів з великою точністю (2—3°) здатні визначати положення джерела звуку. Особливо висока точність звукової локації (близько 1°) у сов, які успішно ловлять здобич «на слух», не бачачи її. У деяких птахів, що гніздяться в глибоких темних печерах, виявлена звукова локація: видаючи в чутному, а не ультразвуковому, як кажани, діапазоні (1,5-7 кгц) уривчасті звуки й уловлюючи їхнє відбиття, птахи в темряві облітають перешкоди й знаходять своє гніздо.
Гострий слух і здатність до акустичного аналізу сполучається в птахів зі здатністю видавати різноманітні звуки, що несуть ємну й біологічно важливу інформацію (звуки спілкування батьків і пташенят, сигнали тривоги, звуки й пози, що регулюють територіальні відносини, взаємини партнерів і т.д.). Голос і слух забезпечують можливості звукового спілкування не тільки між особинами свого виду, але й між різними видами. Так, наприклад, звук тривоги, виданий у лісі сойкою або сорокою, сприймається як сигнал, небезпеки птахами багатьох видів. Подібні по зовнішньому вигляді близькі види звичайно легко розрізняються по пісням і позивам. Тому голос відіграє важливу роль ізолюючого механізму, що попереджає міжвидові схрещування. Надбання стійких відмінностей у позивах і піснях у різних популяцій одного виду підсилює ізоляцію (роз'єднання) цих популяцій і в остаточному підсумку може призвести до їхньої репродуктивної ізоляції, тобто до появи нових видів. Запис сигналів тривоги або страху та наступне їхнє відтворення дозволяє лякати птахів від тих місць, де вони в цей момент можуть заподіювати шкоди (наприклад, шпаків від вишневих садів і виноградників у період дозрівання ягід, багатьох птахів від аеродромів і т.п.).
Основним звуковідтворюючим органом птахів служить нижня гортань; трахея резонує (підсилює) звуки, а верхня гортань, можливо, їх трохи модифікує. Деякі птахи здатні видавати так називані інструментальні звуки: клацання дзьобом, свист повітря в махових (качки, стрепети й ін.) або кермових (бекаси) пір'ях у польоті, удари крильми (голуби, дрімлюги) і т.п. Дятли видають голосну барабанну дріб, ударяючи клюром об сухий сучок. Ці сигнали також несуть біологічну інформацію й служать для спілкування.
Зазвичай вважають, що нюх у птахів розвинено слабко. При цьому ґрунтуються на тому, що майже у всіх птахів нюхові частки переднього мозку малі. Однак у порівнянні із плазунами в птахів збільшується поверхня носової порожнини завдяки розростанню носових раковин і зростає площа нюхового епітелію. Ці факти й дані поки нечисленних експериментів дозволяють припускати, що в деяких птахів (що харчуються падлом хижаки, кулики, качки, імовірно, дятли, деякі горобині й ін.) нюх розвинений досить добре й використається при пошуках їжі. Так, качки вологий звичний корм виявляють по запаху на відстані до 1,5 м. У новозеландських ківі нюх служить провідним рецептором при пошуках корму (до речі, очі в ківі відносно дуже малі).
Смакові бруньки (скупчення аналізуючих клітин, які обплетені нервовими закінченнями) розташовані в слизовій оболонці ротової порожнини, на язиці й у його підстави. Смак у птахів розвинений: багато видів розпізнають солодке, солоне, гірке. У шкірі місцями численні вільні закінчення нервів, що відчувають, або більш складні органи шкірного чуття, що представляють собою скупчення клітин, що відчувають, обплетеними нервовими закінченнями, і опорних клітин. Вони виявлені на оперених і безперих ділянках шкіри, на дзьобі й у ротовій порожнині, на задніх кінцівках. Деякі із цих утворень виконують функцію термодетекторів (реєструють зміни температури тіла), інші несуть функцію рецепторів дотику (зокрема, реагують і на зміну положення пір'я). У видів, що добувають корм у м'якому ґрунті (кулики, гусеподібні, фламінго, удоди й ін.), численні тактильні тільця розташовані в спеціальних дрібних поглибленнях на дзьобі й допомагають у виявленні здобичі.
Поводження й спосіб життя
Загальні особливості поведінки. Прогресивний розвиток центральної нервової системи й органів чуттів, високий рівень обміну речовин і теплокровність забезпечили птахам у порівнянні з плазунами значно більшу рухливість, розширили сприйняття навколишнього світу й ускладнили поводження. Чіткіше проявляються елементи активного пристосування середовища до своїх потреб - гніздобудівнициво, запас їжі, групові ночівлі.
Основу поводження птахів становлять складні комплекси спадково закріплених уроджених (безумовних) рефлексів, що визначають найважливіші елементи їхнього життя: зустріч статі, гніздобудівництво, насиджування й вигодовування молодняку, прийоми, видобутки їжі, міграції й багато чого іншого. Як безумовні подразники виступають окремі елементи середовища (місце для гнізда й підходящий будівельний матеріал при гніздобудівництві, укорочення дня й погіршення кормових можливостей при міграціях і т.п.) і інші особливості свого виду (їхньої пози, рухи й елементи й т.д.). Але поводження кожного птаха істотно збагачується й удосконалюється шляхом надбання індивідуального досвіду (тобто вироблення умовних рефлексів). Наслідування поводження батьків або партнера зграї полегшує вироблення тимчасових зв'язків і збільшує можливості доцільного поводження при змінах зовнішньої обстановки - появі нових кормів, нових джерел небезпеки й т.п. Завдяки цьому досвід однієї особини стає досвідом популяції. Все це збільшує виживаня.
У птахів встановлена наявність «екстраполяційних рефлексів» - здатність вгадувати найближчий розвиток подій (Л. В. Крушинский). Так, при наближенні автомобіля багато птахів злітають із дороги на узбіччя й після цього не реагують на проїжджаючу машину; таким чином, птах оцінює, де пройде машина. Здобич, що прошмигнула у кущ, хижак нерідко очікує з протилежної сторони куща. Таке поводження може розглядатися, як прояв елементів розумової діяльності, не вираженої в плазунів. Екстраполяційні рефлекси краще розвинені у видів, що харчуються рухливою здобичу, - воронових, хижаків й ін.
Птахам властиві афекти - стан страху, злості, радості, спокою, яким відповідає й певне зовнішнє вираження: пози, положення оперення, видавані звуки. Безсумнівна наявність тривалої пам'яті (папуга впізнав свою господарку через 19 років). Здатні птахи й до асоціацій. Наприклад, ворони відрізняють мисливця з рушницею від людини із ціпком і т.д.
Популяційна організація птахів значно складніша й різноманітніша, ніж у плазуныв. Вона міняється по сезонах року. Прихильність до певної території особливо чітко виражена в сезон розмноження. Звичайно птахи, що гніздяться поодиноко, займають певну ділянку, що безпосередньо примикає до гнізда (гніздова територія), на яку не допускають інших особини свого виду. Якщо корм збирається вдалині від гнізда, то площа охоронюваної території буде невелика. У багатьох видів, особливо дрібних горобиних птахів, гніздова територія збігається з кормовою ділянкою й енергійно захищається. Сутички на границі охоронюваної території носять у значній мірі турнірний характер. Практично завжди перемагають хазяї ділянки (здійснюється право першого, а не більш сильного). Завдяки цьому популяція більш-менш рівномірно заселяє придатну територію, ефективно використовуючи її харчові ресурси.
Частина птахів, здатних добувати корм удалині від гнізда, гніздяться колоніями (трубконосы, голінасты, з горобиних — граки, шпаки, ластівки й ін.). При цьому розміри території, охоронюваної від сусідніх особин, часто рівні лише тій відстані, на яке птах, що насиджує, може витягнути дзьоб. Розміри колоній варіюють від деяких десятків пар до десятків тисяч пар, що гніздяться. Часто утворюються змішані колонії, у яких кожен вид займає найбільш для нього зручні ділянки. Розташовані по північних морських узбережжях так звані пташині базари звичайно утворені декількома видами. На широких скелястих уступах щільними групами гніздяться кайри, відкладаючи свої дйца на скелю без усякого гнізда. На дрібних уступах із гниючих водоростей будують об'ємисті гнізда чайки мойовки. У вузьких ущелинах, частіше в нижній частині обриву гніздяться чистики, а в торфі, що покриває скелі, риють гніздові нори тупики. Ці види є основною частиною населення пташиних базарів Європейської Півночі.
Колоніальне гніздування дозволяє із граничною ефективністю використовувати наявні харчові ресурси й придатні для гніздування ділянки, а також забезпечує більшу безпеку членів колонії, тому що деякі хижаки не насмілюються наближатися до таких скупчень, а інших активно відганяють спільними атаками члени колонії. Наприклад, чайки й крачки успішно відганяють від колонії навіть таких хижаків, як лисиць й песців. По відношенню один до одного члени колонії проявляють знижену агресивність, реагують на тривожні сигнали особин свого й іншого видів.
Після закінчення періоду розмноження лише деякі дорослі птахи залишаються на своїх гніздових ділянках до наступного періоду розмноження (ворони, великі строкаті дятли й ін.). Більшість видів міняє осілий спосіб життя на кочовий. У частини видів, яких називають осілими, ці кочівлі обмежуються невеликою територією (переміщення на кілометри, рідше десятки кілометрів), в інших довжина кочівель може становити сотні й тисячі кілометрів (останню групу видів називають перелітними). Переміщення дозволяють птахам вибирати більш кормові ділянки, використовувати кращі укриття від непогоди при ночівлях.
Поза періодом розмноження деякі види тримаються поодинці або парами, більша частина видів поєднується в групи або зграї, що включають десятки й сотні особин (качки, граки, шпаки й багато хто інші). У частини видів родини зберігають у зграях відособленість (гусаки, журавлі), в інших при утворенні зграй родини розпадаються. У зграях звичайно виникає певна організація, заснована на відносинах домінування — підпорядкування, що запобігає сутички між членами зграї й забезпечує їхнє нормальне поводження. Іноді утворюються змішані зграї, що складаються з особин декількох видів. Наприклад, восени й узимку в наших лісах нерідко можно зустріти змішані зграйки, що кочують, з декількох видів птахі. Зграйний спосіб життя полегшує пошук їжі (одночасно обстежується більша територія) і дозволяє легше й швидше виявити небезпеку.
Річні цикли. Сезонна зміна умов життя (погода, їжа) у більшості районів світу визначає річну ритміку зміни стану організму (у тому числі рівня й характеру метаболізма), поводження й популяційної організації птахів. Обумовлююча цю ритміку перебудова гормональної системи здійснює по сигналах зовнішнього середовища. У помірних і високих широтах як такий сигнал першорядне значення має світловий режим (зміна довжини дня й ночі), у тропіках - чергування сухих і вологих періодів. Важливими, але додатковими сигналами служит загальний хід погодніх умов, кількість й якість доступних кормів. Строки, тривалість і характер прояву окремих фаз річного циклу різні в різних груп птахів і визначаються кліматичними особливостями заселених ними районів, характером займаючих місць і видовими екологічними особливостями. Можна виділити наступні основні періоди річного циклу.
Підготовка до розмноження. Початок розвитку статевих залоз під впливом тривалості дня, що збільшується. Переміщення до місць розмноження з місць зимівель, у деяких – формування пар вже на зимівлях або під час перельоту. У частини видав - завершення передстатевої ліньки, що почалася на зимівлі.
Розмноження. Заняття гніздових ділянок, токові явища, формування пар, дозрівання статевих клітин, будівля гнізд, відкладання яєць, їх насиджування й вигодовування пташенят. Він завершується, коли повністю оперені й набули здатність до польоту молоді птахи починають самостійне життя, часто поєднуючись у зграї. У цих зграях можуть бути дорослі й молоді, але зв'язок між пташенятами і їх батьками зазвичай обривається, виключення: гусаки, лебеді, журавлі й ін.
Післягніздове линяння. У птахів після розмноження йде повна післягніздова лінька, коли змінюється все оперення. У полігамних видів самці, що не беруть участь у насиджуванні, починають линяння зразу після завершення яйцекладки. Самці, що линяють, поодинці тримаються в глухих ділянках лісу, а самці качок скупичуються на сильно зарослих озерах іноді за десятки й сотні кілометрів від місць розмноження. Самки починають линяти пізніше, коли пташенята вже підростуть; у них період линяння накладається на кінець періода розмноження. Так само чітко не розмежовані за часом кінець розмноження й початок ліньки в моногамних птахів, дорослі починають ліньку в період завершення годівлі пташенят (імматуронатні птахи) або (у матуронатних видів) коли пташенята підрастуть і стануть більш самостійними. Завершення линяння в частини видів закінчується лише на зимівлі.
4. Період підготовки до зими. Широкі кочівлі в пошуках корму, інтенсивне харчування. Змінюється характер обміну й відбувається поси- ленне накопичення жиру. У пошуках корму деякі види відвідують біотопи, у яких протягом року вони не бувають. Качки й гусаки годуються на хлібних полях, журавлі поїдають картошку, що залишилася. На полях годуються дрозди, голуби, тетерева й інші лісові птахи.
У цей період деякі види птахів роблять запаси корму. Кедрівки— Nucifraga caryocatactes викльовують із шишок горішки кедра й закопують їх у мохи, ховають між каменями й коріннями іноді на відстані декількох кілометрів від кедровників. Частина запасів потім використовується самими птахами, частину поїдають гризуни й комахи, частина насінь проростає. Природне поновлення кедра йде практично тільки таким шляхом. Сойки запасають жолуді дуба, горішки бука, на полях, що примикають до лісу, збирають і ховають у лісі дрібну картоплю. Свій запас сойки розшукують і використовують протягом всієї зими. За рахунок проростання вцілілих запасів жолудів й горішків іде природне поновлення дуба й бука на горах і вирубках.
Повзики ховають горішки бука, насіння клена, в'яза, липи в тріщинах кори. Горобиний і мохноногий сичі — Glaucidium passerinum й Aegolius funereus восени ховають у дуплах і штучних гніздах трупи гризунів. В одному дуплі іноді буває до 50- 80 трупів. Ці запаси використовуються взимку, коли випав сніг ускладнює вилов звірків. У цих випадках запасами користується та ж особина, що і сховала їх. Виявленню запасів, імовірно, допомагає пам'ять і нюх. В останні роки було встановлено, що зграйки синиць, які кочують восени, продовжують пошуки корму й ховають дрібні насіння, ягоди ялівця, лялечок комах у тріщини кори, під нарости лишайника на стовбурах і гілках. Ці запаси використовуються в період зими, при недостачі харчів, іншими особинами цих видів. На відміну від ссавців серед птахів не має видів, які б повністю задовольняли свою потребу в їжі взимку тільки за рахунок запасів; але запас кормів полегшує зимівлю.
Цей період річного циклу завершується переміщеннями птахів на місця зимівель; ці переміщення описуються нижче.
5 . Зимівля.
Популяції кожного виду розміщаються
в районах, що
забезпечує їм їжу й
захист. Частіше в районі зимівель птахи
роблять невеликі кочівлі; у частини
видів (різні гусеодібні, деякі горобині)
чітко виражені добові переміщення з
місць годівлі на місця відпочинку й
назад. Багато морських птахів у цей
період широко кочують по океані,
розшукуючи скупчення корму (трубконосі,
чистикові). Границю між зимівлями й
наступним
періодом
річного циклу - підготовкою до розмноження
- провести важко: у частини видів ще на
зимівлях починається предшлюбне
линяння, іде формування пар, починають
проявлятися токові явища; зимівельні
кочівлі поступово переростають у
спрямовану міграцію до місць гніздування.
. Зимівля.
Популяції кожного виду розміщаються
в районах, що
забезпечує їм їжу й
захист. Частіше в районі зимівель птахи
роблять невеликі кочівлі; у частини
видів (різні гусеодібні, деякі горобині)
чітко виражені добові переміщення з
місць годівлі на місця відпочинку й
назад. Багато морських птахів у цей
період широко кочують по океані,
розшукуючи скупчення корму (трубконосі,
чистикові). Границю між зимівлями й
наступним
періодом
річного циклу - підготовкою до розмноження
- провести важко: у частини видів ще на
зимівлях починається предшлюбне
линяння, іде формування пар, починають
проявлятися токові явища; зимівельні
кочівлі поступово переростають у
спрямовану міграцію до місць гніздування.
Практично всі плазуни й багато ссавців на несприятливі сезонні зміни умов життя відповідають однозначно — зниженням активності й анабіозом. Птахам це не властиво. Однак американські дрімлюги — Рhаlаепорtilus пиttаlliі впадають у дійсну спялчку, що триває 2—2,5 місяці: у птаха, що забилося в ущелину (мал. 62) температура тіла знижується до 18-19º , сповільнюється подих і пульс. Короткочасне заціпеніння при різких спадах температури повітря відзначені й в інших дрімлюг, у стрижів і ластівок. Нічне заціпеніння характерно для багатьох видів колібрі. За цими нечисленними виключеннями птахи не знижують активності при сезонних несприятливих змінах і переживають їх, змінюючи місцеперебування й переходячи на доступні, хоча й менш калорійні корми, змінюючи поведінку (ночівлі в снігу тетерячих птахів і синиць, групові ночівлі в притулках горобців, пищух й ін.), або, летячи на великі відстані, протягом усього року живуть при відносно сприятливих погодних і кормових умовах.
По відношенню до території птахів можна розділити на три групи: 1) осілих - залишаються в тому самому районі протягом усього року; вони можуть весь рік триматися на своїй гніздовій ділянці або міняють місцеперебування, але довжина переміщень звичайно не перевищує кількох десятків кілометрів; 2) що кочують - здійснюють після сезону розмноження ненаправлені кочівлі довжиною сотні кілометрів, але звичайно що не вилітають за межі тієї природної зони, у якій вони гніздяться; 3) перелітних - летять на зимівлі за тисячі кілометрів від місць гніздування, в інші природні зони.
Розподіл видів по цих групах ускладнено тим, що популяції одного виду з різних частин ареалу можуть поводитися неоднаково. Так, сірі ворони в південних районах європейської частини СРСР осілі, у центральних районах кочуючі, а в північних районах дійсні перелітні птахи. Позначаються на характері рухливості птахів і зміна погодних і кормових умов по роках. Дрозди рябинники - перелітні птахи, але в теплі зими, в роки врожаю горобини і ялівця, великі зграї всю зиму кочують у середній смузі. Чим різкіше сезонні зміни умов життя, тим менше загальне число видів, що зустрічається тут, і тим більше серед них перелітних.
Осілі або кочуючі у невеликих межах курячі, крім тундрових популяцій білої куріпки, що відлітають на зиму в лісову зону, і перелітних перепелів. У багатьох районах СРСР осілі дятли, синиці, багато воронових, горобці, але в північних районах вони кочуючі і навіть перелітні. До типово кочуючих видів відносяться: омелюхи, клести, снігурі, чечітки, багато сов й ін. Більшість або всі популяції, приблизно 600 видів з 750, що зустрічаються на території нашої країни, перелітні.
Відносно невелике число видів й особин гусеподібних, поганок, голенастих, хижаків, куликів, чайок, горобиних зимують у південних районах СРСР по берегах Чорного моря, у Закавказзі, на півдні Каспію, у деяких районах Середньої Азії. Переважна більшість видів й особин наших птахів зимує за межами країни на Британських островах й у Південній Європі, у Середземномор'ї, у багатьох районах Африки й Азії. Наприклад, у Південній Африці зимує багато дрібних птахів з європейської частини СРСР, що пролітають від місць зимівель до 9—10 тис. км. Пролітні шляхи деяких видів ще довщі. Полярні крачки — Sterna paradнseo, що гніздяться по узбережжях Баренцева моря зимують на узбережжі Австралії, пролітаючи лише в одну сторону до 16—18 тис. км. Майже такий же пролітний шлях у сивок бурокрылых — Charadrius dominica, що гніздяться в тундрах Сибіру, які зимують у Новій Зеландії, і в колючехвостих стрижів — Hirundapus caudacutus, зі Східного Сибіру, що відлітають в Австралію й Тасманію (12—14 тис. км); частину шляху вони пролітають над морем.
Під час міграцій птахи летять зі звичайними швидкостями, чергуючи переліт із зупинками для відпочинку і годівлі. Осінні міграції звичайно відбуваються з меншою швидкістю, ніж весняні. Дрібні горобині птахи при міграціях за добу переміщаються в середньому на 50—100 км, качки — на 100—500 км і т.п. Таким чином, у середньому за добу птахи витрачають на переліт відносно невеликий час, іноді всього лише 1—2 год. Однак деякі навіть дрібні наземні птахи, наприклад американські деревні славки — Dendroica, мігруючи над океаном, здатні пролітати без зупинки 3—4 тис. км. за 60—70 год. безперервного польоту. Але такі напружені міграції виявлені лише в невеликого числа видів.
Висота польоту залежить від багатьох факторів: виду птаха і її летних здатностей, погоди, швидкості повітряних потоків на різній висоті і т.п. Спостереженнями з літаків і за допомогою радарів було встановлено, що міграції більшості видів проходять на висоті 450-750 м; окремі зграї можуть пролітати й зовсім низько над землею. Значно рідше пролітних журавлів, гусаків, куликів, голубів відзначали на висотах до 1,5 км і вище. У горах зграї куликів, гусаків, журавлів, що летять, відзначали навіть на висоті 6-9 км над рівнем моря (на 9-му кілометрі вміст кисню на 70% менше, ніж на рівні моря). Водні птахи (гагари, поганки, чистикові) частина пролітного шляху пропливають, а деркач проходить пішки. Багато видів птахів, звичайно активні тільки в денний час, мігрують в ночі, а вдень годуються, інші й у період міграції зберігають звичайну добову ритміку активності.
У перелітних птахів у період підготовки до міграцій змінюється характер обміну речовин, що призводить при посиленому харчуванні до накопичення значних жирових запасів. При окислюванні жири виділяють майже вдвічі більше енергії, чим вуглеводи й білки. Резервний жир у міру потреби надходить у кров і доставляється в працюючі м'язи. При окислюванні жирів утворюється вода, чим компенсується втрата вологи при подиху. Особливо великі запаси жиру у видів, змушених під час міграції тривалий час летіти без зупинок. У вже згадуваних американських деревних славок перед польотом над морем запаси жиру можуть становити до 30-35% їхньої маси. Після такого «кидка» птахи посилено годуються, відновлюючи енергетичні резерви, і знову продовжують переліт.
Зміна характеру обміну, що готує організм до перельоту або до умов зимівлі, забезпечується сполученням внутрішньої річної ритміки фізіологічних процесів і сезонних змін умов життя, у першу чергу зміною довжини світлового дня (подовженням - навесні й укороченням - наприкінці літа); імовірно, певне значення має й сезонна зміна кормів. У птахів, що накопили енергетичні ресурси під впливом зовнішніх стимулів (зміна довжини дня, погода, недолік кормів) наступає так зване «перелітне занепокоєння», коли поводження птаха різко міняється й виникає прагнення до міграції.
У переважної більшості кочуючих і перелітних птахів чітко виражений гніздовий консерватизм. Він проявляється в тому, що птахи на наступний рік повертаються із зимівлі на місце попереднього гніздування і займають старе гніздо, або поблизу будують нове. Молоді птаха, що досягли статевої зрілості повертаються на свою батьківщину, але частіше поселяються на якійсь відстані (сотні метрів - десятки кілометрів) від того місця, де вони вилупилися (мал. 63). Менш чітко виражений у молодих птахів гніздовий консерватизм дозволяє виду заселяти нові, придатні для нього території й, забезпечуючи перемішування популяції, запобігає інбридінгу (близькородне схрещування). Гніздовий консерватизм дорослих птахів дозволяє їм гніздитися в добре знайомому районі, що полегшує і пошук їжі, і порятунок від ворогів. Існує й сталість місць зимівель.
Як птахи орієнтуються під час міграцій, як вибирають напрямок перельоту, потрапляючи в певний район на зимівлю й повертаючись за тисячі кілометрів на місце гніздування? Незважаючи на різноманітні дослідження, відповіді на це питання поки немає. Мабуть, у перелітних птахів є вроджений міграційний інстинкт, що дозволяє їм вибирати потрібний загальний напрямок міграції. Однак цей уроджений інстинкт під впливом умов середовища, може швидко змінюватися. Яйця осілих англійських крижнів були інкубовані у Фінляндії. Вирослі молоді крижні, як і місцеві качки, восени полетіли на зимівлю, а наступною весною значна їхня частина (36 з 66) повернулася у Фінляндію в район випуску й там загніздилася. В Англії жодна із цих птахів не була виявлена. Чорні казарки перелітні. Їхні яйця інкубувались в Англії, і молоді птахи восени поводилися на новому місці як осілі птахи. Таким чином, пояснити й саме прагнення до міграції, і орієнтування під час перельоту тільки вродженими рефлексами поки не можна.
Експериментальні дослідження й польові спостереження свідчать, що мігруючі птахи здатні до астронавігації: до вибору потрібного напрямку перельоту по положенню сонця, місяця й зірок. При похмурій погоді або при зміні картини зоряного неба при дослідах у планетарії здатність до орієнтації помітно погіршувалася.
Здатність до астронавігації припускає наявність «біологічних годинників», які дозволяють ураховувати зміну положення небесних орієнтирів протягом доби. Експерименти показали, що почуття часу в птахів має точність 10-15 хв. Цього досить, щоб вибрати правильний напрямок польоту. Ряд експериментів і спостережень дозволяє припускати, що птахам властиво «компасне відчуття» - здатність визначати правильний напрямок при перельотах або при завезенні далеко від гнізда; воно може проявлятися й у похмуру погоду, коли астронавігація затруднена. Широкий розвиток
Рис. 63. Розселення молодняку сипух (Туtо) за даними кільцювання (по Стюарту).
Цифри на колі - відстань від місця народження (у милях), цифри між колами - число виловлених окільцьованих птахів
досліджень дозволяє сподіватися, що найближчим часом будуть розкриті механізми, що визначають високі можливості птахів до орієнтування в просторі і їхні навігаційні здатності.
Обраний правильний загальний напрямок польоту коректується зорово, тому що птахи при міграціях дотримуються звичних ландшафтів — русел рік, лісових масивів і т.п. При перельотах змішаними за віком зграями орієнтування полегшує досвід особин, що вже робили міграції. Однак у великої кількості видів молоді птахи летять не разом з дорослими, а самостійно, раніше (багато горобиних, деяких хижаків й ін.) або пізніше (багато куликів, деякі горобині) старих птахів. Імовірно, наступне повернення на місця гніздування полегшується гарним знайомством з місцевістю під час післягніздових кочівель, які в різному ступені виражені у всіх видів.
У більшості птахів те або інше відношення до території складалося одночасно зі становленням виду, тому що зміни пір року в багатьох районах земної кулі були виражені вже в крейдовому — третинних періодах — часу становлення сучасних груп. Великі площі, що захопили інтенсивні процеси горотворення й посилення контрастності клімату, кілька заледенінь, що формувалися в багатьох районах Північної Америки і Євразії в четвертинний період, імовірно, підсилили рухливість птахів у позагніздовий час. Після танення льодовиків птахи інтенсивно заселяли території, що звільнилися. Поступово формувалися сучасні зимівлі й напрямки прольоту до них. Цей процес триває й у цей час. Створення водоймищ по трасі Каракумского каналу супроводжувалося виникненням нових зимівель водних птахів. З іншого боку, інтенсивне господарське використання багатьох районів півдня Європи привело до порушення природних ландшафтів і до різкого зниження числа зимуючих там птахів.
Вивченню міграцій птахів допомогло масове їхнє кільцювання: молодому або виловленому дорослому птаху на ногу одягають легке металеве кільце з номером й умовною позначкою установи, що провела кільцювання. Кільцювання проводять багато наукових установ. Всі відомості про окільцьованих тварин надходять у Центр кільцювання Академії наук СРСР (Москва). Туди ж потрібно посилати кільця, зняті з добутих птахів. Щорічно в усім світі кільцюють близько 1 млн. птахів, у тому числі в СРСР - приблизно 100 тис. Масове кільцювання дозволило з'ясувати для ряду видів ступінь гніздового консерватизму, напрямки й швидкість міграцій, місця зимівель, тривалість життя й зразкові розміри смертності, сталість пар і т.п.
Поширення птахів й їхня роль у біоценозах. Птахи поширені по всій земній кулі: вони зустрічаються на всіх континентах, включаючи великі пустелі й суворі високогір'я, на самих віддалених океанічних островах й у центральних районах океанів за тисячі кілометрів від найближчих ділянок суши. Навіть у полярні ночі на розводдях серед льодів Льодовитого океану зустрічаються, хоча й у невеликих кількостях, деякі птахи (білі чайки — Pagophila ebиrnea, кайри, люрики — Plotus alle й ін.). Настільки широке поширення забезпечується всім комплексом характерних для птахів особливостей - великою рухливістю (політ), гомойотермністю, високим рівнем вищої нервової діяльності, різноманіттям морфо-фізіологічних адаптації й т.п.
Більшість видів птахів — близько 80% — знаходяться у тропічній зоні; до півночі й до півдня число видів, що зустрічаються, зменшується. У Полеарктиці, що становить 34% всієї площі суши земної кулі, зустрічається 1100 видів птахів, в Африці (близько 20% суши) — 1700 видів, а в Південній Америці, територія якої становить лише близько 12% суши, але лежить переважно в зоні тропіків, — 2600 видів. Найбільш різноманітний видовий склад птахів у лісах, найбільш бідний - у пустелях і тундрах. Так, у Тиманській тундрі (Архангельська обл.) і в Центральних Каракумах гніздиться приблизно однакове число видів птахів - близько 60, але за рахунок пролітних і зимуючих птахів число видів, що зустрічаються в Каракумах, зростає до 220. У районах, де добре виражена мозаїчність ландшафтів (водойми в лісах або пустелях, стики лісу й лугів і т.п.), зустрічається більше число видів, чим в однотипних ландшафтах того ж географічного району.
Кожний вид пристосований до життя в одному або декількох ландшафтних співтовариствах - біогеоценозах. Ця пристосованість виражається в особливостях руху, наборі використовуваних кормів і способах їхнього добування, стійкості до характеру клімату, ступеня осілості, загальному характері поведінки й т.п. По ландшафтних зв'язках можна виділити кілька великих екологічних груп птахів.
Найбільш численна по числу видів група деревинно-чагарникових птахів, що знаходяться в лісах різних типів і чагарникових заростей (див. мал. 37). Серед наших птахів цієї групи є тваринноїдні, всеїдні й рослинноїдні. У тропіках досить багато дрібних птахів, що п'ють нектар і поїдають тичинки і пильники; вони забезпечують перехресне запилення багатьох рослин. Більшість видів годується на гілках, рідше - на землі. Гнізда звичайно розміщаються в розвилках гілок, у частини видів - у дуплах. Найбільш чітко деревна спеціалізація проявляється в папуг, які спритно лазают у гілках за допомогою задніх кінцівок і дзьоба, і в дятлів, що вільно пересуваються по вертикальних стовбурах, чіпляючись лабетами й опираючись на твердий хвіст, і долблять кору й деревину в пошуках комах. Деякі деревні птахи годуються більше на землі, іноді вилітаючи на годівлю на луги й поля (шпаки, дрозди, голуби й ін.). Лісові тетерячі (глухар, рябчик й ін.) гніздяться на землі й у безсніжний період годуються теж на землі, а взимку годуються в кронах, поїдаючи: глухарь - хвою, а рябчик - бруньки й сережки.
Птахи відкритих просторів - лугів, степів, пустель (див. мал. 36) - гніздяться й годуються на землі. Цю групу становлять страуси, дрохви, деякі кулики, рябки, з горобиних сюди можна віднести жайворонків, ковзанів, каменок (останні будують гнізда під каменями й у норах гризунів). По плямах чагарникових заростей у ці ландшафти проникають і деревинно-чагарникові птахи.
Болотні птахи населяють заболочені луги, осокові й мохові болота, зарості по берегах водойм. Здобич звичайно збирають на землі. Великі довгоногі й довгошиї види (журавлі, чаплі, лелеки) легко ходять, переступаючи через купини й густу траву і схоплюють здобич, а дрібні, теж часто довгоногі види (дрібні чаплі, пастушки, багато куликів), спритно бігають у густій траві; прослизаючи між стеблами. До цієї групи близькі й багато інших куликів, що звичайно годуються на обмілинах. На купинах і стеблах улаштовують гнізда й горобині птахи - очеретники, цвіркуни й ін.
Птахів, що повністю перейшли до водного способу життя й втратили всякі зв'язки із сушею, немає: всі водні птахи в тім або іншому ступені зберігають зв'язок із сушею й повітрям (див. мал. 35). Загальними пристосуваннями до водного способу життя будуть щільне оперення з добре розвиненим пухом, плавальні перетинки, що з'єднують пальці задніх кінцівок, швидкий, але не маневрений політ (пінгвіни втратили здатність до польоту) і т.п. Гнізда влаштовують по берегах водойм на землі й скелях, заломах очерету, на деревах, у норах й ущелинах скель. Гагари, поганки, баклани, чистикові й пінгвіни їжу добувають тільки у воді, плаваючи й пірнаючи. По суші пересуваються мало й незграбно. Тільки у воді добувають їжу - рибу - і пелікани, але пірнати вони не можуть. Частина гусеподібних годується тільки у воді, плаваючи й пірнаючи, річкові качки годуються на мілководдях, занурюючи у воду голову й передню частину тулуба, і на суші. Гусаки годуються переважно на суші, поїдаючи зелені пагони й насіння. Чайки, крачки, трубконосі й частина веслоногих (олуши, фрегати, фаетони) мають маневрений політ і можуть перебувати в повітрі багато годин. Добре плавають. Здобич - різноманітних водних тварин - виглядають у польоті й схоплюють у стрімкому пікіруванні, пірнаючи у воду лише на невелику глибину. Крачки здатні скльовувати на льоту здобич з надводних рослин і з землі, а чайки нерідко полюють на мілководдях і на суші «пішки» (ходять вони добре).
У птахів, як й в інших хребетних, добре виражена сезонна динаміка чисельності. Із всіх періодів року найбільш рівномірний розподіл по території, буде в гніздовий період (у наших широтах наприкінці весни — початку літа). У лісах, чагарникових лугах загальна чисельність у середньому дорівнює 4—7 птахів на 1 га, а в найбільш сприятливих ділянках — опушки лісу із сильно розвинутим чагарниковим ярусом і т.п. — може доходити до 50—70 особин всіх видів на 1 га, але така висока чисельність зустрічається рідко. Однак у колоніальних птахів у районі колоній чисельність може бути набагато вище. Так, у червоноклювого ткачика — Q. quelea в саванах Африки колонії можуть складатися з декількох сотень тисяч гнізд, причому на 1 га може розміщатися до 12 тис. гнізд, тобто 24 тис. дорослих птахів; на територіях між колоніями чисельність цих видів буде набагато менше.
На підставі екстраполяції обліків птахів у різних ландшафтах визначають зразкову чисельність всіх видів птахів Північної Америки в період розмноження в 5-6 млрд. Після вильоту молодих загальна чисельність зростає приблизно до 20 млрд. й, поступово знижуючись за рахунок смертності молодих і дорослих, до початку наступного літа досягає первісного рівня. Думають, що загальна чисельність птахів усієї земної кулі становить приблизно 100 млрд. особин. Ці цифри орієнтовні й дають лише саме загальне уявлення про чисельність цього класу. У помірних і північних широтах сезонна зміна чисельності виражена особливо різко ще й тому, що багато (а в тундрах - майже всі) видів на зиму відлітають. Тому взимку населення птахів у цих районах мінімально, навесні після прильоту статевозрілих особин багатьох видів чисельність різко зростає, далі збільшується в 2-3 рази до кінця літа, до часу появи літнього молодняку, і знижується до мінімуму до зими.
Крім цього, чисельність птахів міняється й по роках. Річні зміни чисельності визначаються в першу чергу коливаннями погодних умов (рання або пізня весна, сухе або вологе літо й т.п.) і змінами кількості і якості доступних кормів, що позначається на розмірах смертності й результатах розмноження. Наприклад, при сильних і тривалих посухах у багатьох районах Казахстану перестають гніздитися водоплавні птахи, а при різкому зниженні чисельності лемінгів у тундрах зменшують розміри кладки або зовсім не приступають до розмноження соколи сапсани й кречети, полярна сова й деякі інші види. Природно, що на наступний рік їхня чисельність істотно знизиться. Ці багаторічні зміни чисельності птахів вивчені ще слабко.
Однак можна сказати, що чисельність птахів по роках міняється у відносно невеликих межах: в 2—4, рідше до 10 разів. Прямий і непрямий вплив людини на фауну приводить до того, що в більш обжитих районах у багатьох видів поряд з багаторічними коливаннями чисельності відбувається їх неухильне зниження. Така справа із чисельністю багатьох видів гусеподібних, журавлів, лелек, хижих птахів. У той же час деякі види, що знайшли оптимальні умови життя в антропогенному ландшафті, поступово збільшують чисельність і помітно розширюють свій ареал. Завезені наприкінці минулого століття кілька десятків шпаків і будинкових горобців успішно розмножилися й розселилися зараз майже по всій Північній Америці, досягши приблизно такої ж чисельності, як на батьківщині. На південно-західній частині СРСР в останні десятиліття розселяється кільчаста горлиця — Streptopelia decaocto, із Закарпаття, що проникнула до ряду пунктів Центрально-Чорноземного району й Прибалтики, по деяких районах Середньої Азії розселяється майна —Acridotheres tristis і т.п.
Відносно висока чисельність птахів, велика рухливість і високий рівень метаболізму визначають їхню видну роль у біогеоценозах. Птахи звичайно представляють кінцеві ланки ланцюгів харчування. Досить значна їхня роль у поширенні насінь ряду рослин (кедр, дуб, горобина, малина й ін.); деякі птахи виконують функцію запильників. Під час кочівель і сезонних міграцій птахи відвідують біоценози, розділені сотнями й тисячами кілометрів, і тим самим беруть участь у глобальному біологічному круговороті речовин.
Значення птахів для людини
Значна роль птахів у житті біосфери обумовлює і їх різноманітне, позитивне й негативне значення для людини.
Винищуючи шкідливих для сільського й лісового господарства тварин (комах, гризунів) і насіння бур'янистих рослин, птахи можуть бути важливим регулятором їхньої чисельності. Розумне використання птахів (у першу чергу шляхом створення достатньої чисельності) може значно знизити необхідність використання хімікатів, неприпустимість широкого застосування яких стає усе більш очевидне. Зараз з'ясовано, що тільки складні лісові насадження з багатим пташиним населенням стійкі до шкідників, тоді як в одновидових, позбавлених чагарникового ярусу лісах (наприклад, соснових) спалаху масової чисельності різноманітних шкідників комах дуже часті й істотно позначаються на якості деревени. Як винищувачі шкідливих комах у лісах особливо корисні всі види синиць, пищуха, повзик, мухоловки, славки, іволги, дрімлюги, зозулі, дятли, у лугах і на полях - жайворонки, шпаки, ластівки, трясогузки, дрібні соколики (боривітра, кобчик), граки, місцями чайки, сірі куріпки, перепели й багато інших птахів. Гризунів інтенсивно винищують не тільки хижі птахи й сови, але й різні воронові птахи, чайки, лелеки й ін. Корисність птахів у великому ступені пов'язана з їхньою здатністю швидко знаходити скупчення їжі - місця масового розмноження шкідників - і концентруватися там. При достатку їжі птахи можуть переходити на невластиві їм корми. Це дозволяє птахам у багатьох випадках успішно придушувати виниклі вогнища масового розмноження шкідників.
Підвищенню чисельності корисних птахів сприяє створення сприятливих умов для їхнього гніздування (куртини кущів у парках і садах, зменшення занепокоєння в період гніздування, знищення здичавілих кішок і т.п.), для осілих птахів - підгодівля взимку. Розважуванням штучних гніздовій можна збільшити чисельність птахів - дуплогніздників (синиці, мухоловки, шпаки й ін.) в 10-25 разів.
Багато птахів (гусеподібні, курячі, деякі кулики й ін.) служать об'єктами спортивного або (зараз лише в деяких районах) промислового полювання. Раціональне ведення мисливського господарства дозволяє одержувати з великих територій додаткову продукцію у вигляді значної кількості смачного м'яса. У деяких районах Середньої Азії дотепер збереглося полювання з ловчими птахами - соколами, яструбами, беркутами. Досвідчені мисливці з гарними беркутами добувають за промисловий сезон до 50-60 лисиць, багато зайців, а іноді й вовків. Спортивне полювання з ловчими птахами зараз одержує досить широке поширення (наприклад, в Угорщині).
Слід зазначити й велике естетичне значення птахів. Завдяки своїй рухливості, спритності й граціозності, дзвінким вигукам і мелодійним пісням вони прикрашають і пожвавлюють ліси й парки, луги й береги водойм. І з цієї сторони птахи заслуговують усілякої охорони.
Місцями деякі види птахів можуть приносити й певний збиток. Корисні винищуванням різноманітних комах шпаки під час післягніздових кочівель можуть заподіювати збиток, роздзьобуючи дозрілі вишні й виноград. Знищуючи ґрунтових комах, граки одночасно викльовують посіяні насіння й висмикують проростки, особливо кукурудзи. У південних районах подекуди серйозний збиток урожаю зернових, особливо просу, і винограду наносять численні горобці — будинковий — Passer domesticus і чорногрудий — P. hi-spaniolensis. У багатьох випадках цей збиток можна різко скоротити, відлякуючи птахів (зокрема, гарний ефект дає трансляція заздалегідь записаних елементів нещастя або сигналів тривоги) або відповідним образом розміщаючи культури (посіви проса в декількох кілометрах від населених пунктів практично не ушкоджуються горобцями). У деяких випадках доводиться вживати заходів по обмеженню чисельності окремих видів. З хижих птахів реальну втрату мисливському господарству може заподіювати лише місцями очеретяний лунь — Circus aeruginosas (і сіра ворона).,
З появою реактивних швидкісних літаків почастішали випадки зіткнення їх із птахами, що іноді приводять до серйозних аварій. Цей збиток попереджається відлякуванням птахів з районів аеродромів і вибором трас польотів в обхід місць сезонних концентрацій птахів.
Поряд з дикими ссавцями птахи забезпечують існування природних вогнищ ряду небезпечних захворювань людини й свійських тварин або як зберігачі й розповсюджувачі збудника, або як годувальники й розповсюджувачі переносників цих збудників (комарів, бліх, кліщів). Роблячи далекі перельоти, птахи сприяють переміщенню збудників навіть між континентами. Зараз установлена роль птахів у циркуляції ряду вірусних захворювань: орнітозів (у тому числі й так називаної хвороби папуг - пситтакоза), грипу, енцефалітів і деяких інших.
Домашні птахи. Птахівництво, що розвивається швидкими темпами, усе більше й більше приймає риси промислового виробництва. На птицефабриках продуктивне поголів'я утримується в клітках; годівля, збір яєць, підтримка певної температури й освітленості механізовано й автоматизовано. У працюючі в автоматичному режимі інкубатори одночасно закладаються десятки тисяч яєць. Висока продуктивність забезпечується й безупинно проведеною селекційною роботою. Одомашнювання птахів було здійснено в далекій давнині. Предком всіх порід домашніх курей були банківскі, або чагарникові, курки — Gallus sp., три види яких і зараз живуть у дикому стані в лісах Індії й Південно-Східної Азії. Дикі кури мають масу 0,7—1 кг, самка відкладає 5—8 яєць. Одомашнені вони були в Індії за кілька тисячоріч до нашої ери, а в Європі з'явилися за 1—2 тисячоріччя до нашої ери. Шляхом селекції зараз виведено близько 100 порід домашніх курей, деякі з них досягають маси 5—7 кг, інші відрізняються дуже високою яйценосністю. Створено й чисто декоративні породи: так, у півнів японських паркових курей довжина хвоста досягає 5—6 м.
Індичка дика — Meleagris gallopavo, що живе в розріджених лісах півдня Північної Америки була приручена мексиканськими індіанцями, імовірно, ще до нашої ери. Сучасні породи відрізняються різноманітним забарвленням, великою масою (індики досягають 12— 15 кг), інтенсивним зростанням молодняку. Різноманітні породи домашніх гусаків — результат одомашнювання й наступної тривалої селекції широко розповсюдженого в Євразії сірого гусака — Anser anser, зустрічається в деяких районах Східної Азії гусак сухонос — A. cygnoides. Різноманітні породи домашніх качок — нащадки широко розповсюдженого в північній півкулі крижня — Anas platyrhunchos (маса 0,8—1,5 кг); деякі породи відрізняються великими розмірами (селезні руанских качок мають масу 4—5 кг), інші — підвищеною яйценосністю (до 200—220 яєць у рік).
Широко розповсюджений у Євразії й у Північній Африці сизий голуб — Columba livia, був приручений одночасно в Індії й у Єгипті приблизно за 5 тис. років до нашої ери. Зараз виведено більше 200 порід, які можна об'єднати в три групи: декоративні, поштові і м'ясні. Декоративні голуби відрізняються своєрідним забарвленням і формою тіла, наявністю пір'я, що прикрашає. До них відносять дутишей, павиних, чубатих, або якобінців, турманів і багатьох інших. М'ясні породи (книг, римські й ін.) відрізняються великими розмірами (маса до 1—1,3 кг, замість 0,3—0,4 кг у дикого голуба). Поштові голуби відрізняються швидким польотом і гарною здатністю до орієнтування. Треновані птахи повертаються на свій голубник з відстані 300—400 км (деякі навіть до 1000 км) зі швидкістю 60—70 км/ч. Голубиний зв'язок використовувся з давніх часів (записка прикріплювалася до ноги) аж до недавнього часу. Лише розвиток радіозв'язку повністю замінив голубину пошту, але в окремих, спеціальних випадках поштові голуби використовуються і зараз.