

X.1. 5. Круговорот железа
В круговороте железа большую роль играют железобактрии. На основании морфологических характеристик все железобактерии могут быть разделены на две группы: нитчатые и одноклеточные. К первой группе относятся грамотрицательные нитчатые бактерии, окруженные чехлом, в которых накапливаются окислы железа и/или марганца (р. Leptothrix, р. Sphaerotilis). Окисление железа (и марганца) и отложение их в чехлах бактерий не связано с получением энергии, идет под действием перекиси водорода. С помощью восстановленных форм железа и марганца обеспечивается удаление Н2О2 – токсического продукта метаболизма:
2Fe2+ + H2O2 + 2H+ 2Fe3+ + 2H2O
Вторая группа бактерий делится на две подгруппы и включает одноклеточные организмы из разных таксонов. Первая подгруппа объединяет железобактерии, растущие в нейтральной или слабощелочной среде и характеризуются хемоорганогетеротрофным типом метаболизма. Сюда относятся свободноживущие микоплазмы р. Metallogenium, р. Gallionella, р. Siderococcus. Окисление железа и/или марганца у данных микроорганизмов – результат химических реакций или фукционирования перекисного пути и не имеет отношения к получению клетками энергии.
Вторую подгруппу составляют в большинстве аэробные ацидофильные формы (основной представитель – Thiobacillus ferrooxidans). Оптимальный рН их роста лежит ниже 4,5 (2–3). Для ацидофильных железобактерий установлена способность получать энергию в результате окисления двухвалентного железа:
2Fe2+ + 1/2O2 + 2H+ 2Fe3+ + H2O
Механизм окисления Fe2+ в дыхательной цепи изучен у Thiobacillus ferrooxidans. Дыхательная цепь этой бактерии содержит все типы переносчиков, характерных для дыхательной цепи аэробных хемоорганофных эубактерий, но участок цепи, связанный с получением энергии, очень короток (рис. 47).
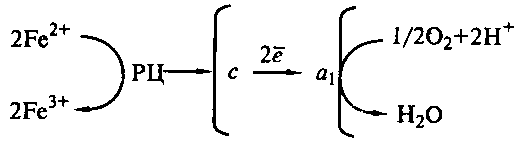
Рис. 47. Схема энергетических процессов у Thiobacillus ferrooxidans: РЦ – рустицианин; c, а1 – цитохромы
Окисление Fe2+ происходит на внешней стороне ЦПМ; в цитозоль через мембрану железо не проникает. Электроны с Fe2+ акцептируются особым медьсодержащим белком – рустицианином, находящимся в периплазматическом пространстве. Затем с рустицианина они передаются на цитохром с, локализованный на внешней стороне ЦПМ, а с него на цитохром а1 , расположенный на внешней стороне мембраны. Перенос электронов с цитохрома а1 на ½ O2, сопровождающийся поглощением из цитоплазмы 2Н+, приводит к восстановлению молекулярного кислорода до H2O. Особенность дыхательной цепи Thiobacillus ferrooxidans – отсутствие переноса через мембрану протонов, а перенос только электронов. Синтез АТФ происходит за счет движения Н+ из внешней среды в цитоплазму через АТФ-синтазный комплекс. Для синтеза одной молекулы АТФ необходимо окислить как минимум 2 молекулы Fe2+ .
Железобактерии широко распространены в природе. Они обнаружены в подземных водах сульфидных месторождений, кислых водах железистых источников и кислых озерах с высоким содержанием закисного железа.
X.2. Типы взаимоотношений микроорганизмов в биоценозах
Благодаря разнообразию механизмов утилизации источников питания и энергии, а также выраженной адаптации к внешним воздействиям, микроорганизмы обитают там, где другие формы жизни не выживают. В зонах обитания микроорганизмы образуют биоценозы (от греч. bios – жизнь + koinos – сообщество) – сложные ассоциации со специфическими и часто необычными взаимоотношениями. Каждое микробное сообщество в конкретном биоценозе образует специфические аутохтонные микроорганизмы (от греч. autos – свой + chthon – страна, местность), т.е. микробы, присущие конкретной области. В состав этих сообществ могут внедряться аллохтонные микробы (от греч. allos – свой + chthon – страна, местность; буквально – чужестранец), обычно в них не встречающиеся.
В природных биоценозах складываются определенные типы взаимоотношений микробов: симбиоз, паразитизм, антогинизм.
Симбиоз (от греч. simbiosis – совместное проживание) – различные формы совместного существования разноименных организмов, составляющих симбиотическую систему. Типичные симбиотические микробы – Escherichia coli, бактерии родов Bacteroides и Bifidobacterum и другие представители кишечной микрофлоы.
Различают несколько форм симбиоза. При мутуализме (от лат. mutuus – взаимный) отношения между партнерами характеризуются взаимовыгодностью и ни один из них не может существовать без другого. Например, микроорганизмы, обитающие в кишечнике человека, вырабатывают биологически активные вещества (витамины группы В), при этом сами защищены от неблагоприятных условий среды (высыхания, экстремальных температур) и имеют постоянный доступ к питательным веществам. Примером мутуализма могут служить также термиты и живущие у них в кишечнике жгутиконосцы, обладающие способностью переваривать поглощаемую термитами клетчатку, которую насекомые без симбионтов не способны усваивать.
При комменсализме (от лат. com – стол, mensa – трапеза; сотрапезничество) выгоду извлекает только один из партнеров. Присутствие комменсала для хозяина остается обычно безразличным и не наносит ему «видимого» вреда. Микроорганизмы комменсалы колонизируют кожные покровы и полости организма человека (например, желудочно-кишечный тракт), не причиняя вреда. Многие бактерии-комменсалы принадлежат к условно-патогенной микрофлоре и способны при определенных обстоятельствах вызывать заболевания (при повреждении кожных покровов).
Паразитизм (от греч. parasitos – нахлебник, от para и sitos – хлеб, пища) форма взаимоотношений двух различных организмов, принадлежащих к разным видам и носящая антагонистический характер, когда один из них использует другого в качестве среды обитания и/или источника пищи.
Паразитические микроорганизмы разделяют на внутриклеточные (вирусы, риккетсии, хламидии) и внеклеточные (большинство бактерий и простейших). В зависимости от внешних условий некоторые микроорганизмы могут вести себя как паразиты либо как сапрофиты. Их называют сапрофитными микроорганизмами, к ним относится большинство условно-патогенных бактерий. Облигатные паразиты полностью утратили собственные метаболитические возможности и живут, разрушая ткани хозяина.
Антагонизм (от греч. antagonizmai – спор, борьба, соперничество) – взаимоотношения при которых один вид микроорганизма задерживает или полностью подавляет рост другого. Антагонистические взаимоотношения особенно выражены в местах естественного обитания большинства видов и типов микроорганизмов, имеющих одинаковые пищевые и энергетические потребности. При этом воздействие на конкурента может быть пассивным или активным. В первом случае микроорганизмы быстрее утилизируют субстрат, лишая соперника «сырьевых ресурсов»; во втором истребляют конкурента различными методами, поглощая их или выделяя высокоспецифичные продукты, токсичные для конкурентов (антибиотики и бактериоцины). Антибиотики (от греч. anti – против + bois – жизнь) – специфические химические вещества, способные в малых количествах оказывать избирательное действие на другие микроорганизмы, подавляя процессы синтеза компонентов клеточной стенки, синтеза белка и нуклеиновых кислот и др. Бактериоциды – белки, синтезируемые отдельными клонами бактерий. В отличие от антибиотиков, секреция бактериоцинов сопровождается гибелью клетки-продуцента. Бактериоциногения (образование бактериоцинов) более выражена у грамотрицательных бактерий, но известна и у грамположительных видов. Известны более 200 различных бактериоционов, обычно обозначаемых по родовому или видовому названию продуцента, – колицины (Escherichia coli), пестицины (Yersinia pestis), стафилоцины (Staphylococcus), вибриоцины (Vibrio). Некоторые бактериоцины действуют на ЦПМ, другие на биосинтез белка.