

Глава X. Экология микроорганизмов
Вездесущность микроорганизмов обусловлена их разнообразием и способностью использовать любые источники питания. В биосфере нет такой среды, в которой не встречались бы микроорганизмы. Образуя сложные ассоциации (микробные сообщества), они играют колоссальную роль в природе, круговороте веществ, поддерживают динамическое равновесие биосферы.
X.1. Биогеохимическая деятельность микроорганизмов
Микроорганизмам принадлежит первостепенная роль в круговороте, миграции, распределении и концентрации химических элементов в биосфере. Некоторые этапы круговорота веществ могут осуществляться только прокариотами.
X. 1. 1. Круговорот углерода
Циклические превращения углерода (и кислорода) реализуются через два разнонаправленных процесса: фотосинтез и дыхание. При фотосинтезе происходит потребление углерода (в виде СО2) и перевод его в органические соединения. В этом процессе участвуют зеленые растения, водоросли и фотосинтезирующие микроорганизмы. Потребление углерода уравновешивается распадом органических веществ. Полное окисление органических веществ до СО2 в присутствии кислорода осуществляют многие аэробные (псевдомонады, бациллы) и факультативно анаэробные бактерии (актиномицеты), грибы, животные. В анаэробных условиях органические соединения расщепляются путем брожения.
Углерод может извлекаться из круговорота. Так, ионы карбоната, содержащиеся в морской воде, соединяясь с растворенными ионами Са2+, осаждаются в виде СаСО3. Карбонат кальция может образовываться и биологическим путем в известковых структурах простейших, кораллов и моллюсков, откладываясь в качестве известняковых горных пород. В условиях высокой влажности и недостатка кислорода отложение отмерших органических остатков приводит к накоплению гумуса, образованию торфа и каменного угля. В результате жизнедеятельности микроорганизмов образуются также метан, нефть.
X.1. 2. Круговорот азота
Цикл азота состоит из нескольких этапов (рис. 43), основную роль в которых играют микроорганизмы, преимущественно бактерии.
Азот составляет 80% земной атмосферы. Как газ азот химически инертен; он не может быть непосредственно использован растениями, животными и большинством микроорганизмов.
Азотофиксация (первый этап) в круговороте азота осуществляется исключительно азотофиксирующими микроорганизмами. Общая продукция микробной фиксации азота составляет до 330 млн т/год (промышленный синтез аммиака из азота и водорода составляет не более 5% фиксированного азота нашей планеты). Азотофиксаторы известны среди 100 родов бактерий, среди которых такие группы, как анаэробные клостридии, сульфатредуцирующие бактерии, энтеробактерии, фотосинтезирующие анаэробы, метанотрофы, спириллы, псевдомонады, актиномицеты, цианобактерии и др.
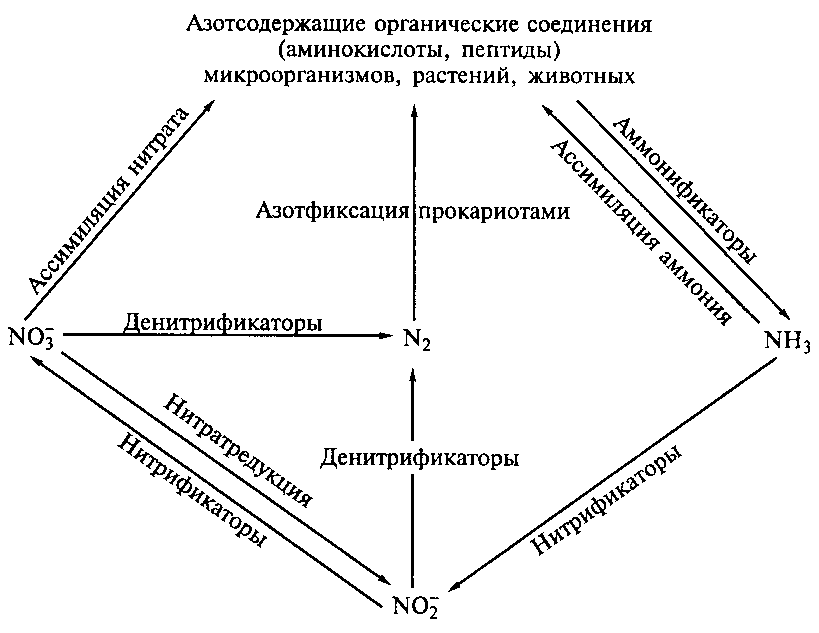
Рис. 43. Цикл азота
Различают симбиотическую и несимбиотическую (ассоциативную) азотофиксацию. Симбиотическая азотофиксация осуществляется бактериями рода Rhizobium (вызывают образование клубеньков у бобовых растений), актиномицетами Franckia (симбионты тропических растений), цианобактериями Anabaena azollae, Nostoc punctiforme.
Несимбиотическая азотофиксация осуществляется бактериями рода Azotobacter, аноксигенными фототрофными бактериями, цианобактериями, клостридиями, факультативными анаэробами Bacillus polymixa, Klebsiella preumonie, хемолитотрофными бактериями Alcaligenes latus, Xanthobacter autotrophicus, метилотрофами, метаногенами и сульфатредуцирующими бактериями.
Фиксация молекулярного азота осуществляется с помощью ферментной системы – нитрогеназы, состоящей из двух компонентов: малого, содержащего железо и серу (Fe-белок), и большого, в состав которого дополнительно входит молибден (МоFe-белок). Соотношение между ними у разных азотофиксирующих микроорганизмов колеблется от 1:1 до 2:1.
Для функционирования нитрогеназы наобходим источник энергии в виде АТФ, ионы магния и восстановитель с низким окислительно-восстановительным потенциалом. Для большинства микроорганизмов непосредственным донором электронов для восстановления азота служит восстановленный ферродоксин, электроны с которого поступают сначала на Fe-белок нитрогеназы (рис. 44). Восстановленный Fe-белок образует комплекс с молекулами Мо и АТФ, что приводит к сдвигу окислительно-восстановительного потенциала FeS-центра белка от –290 до –400 мВ. Это делает возможным перенос активированных электронов на МоFe-белок, в активном центре которого происходит восстановление N2.
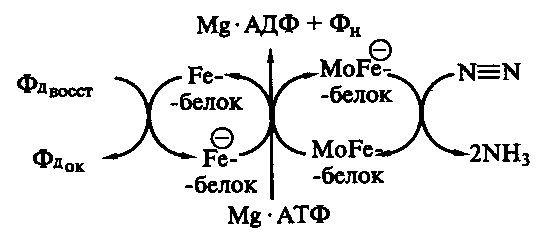
Рис. 44. Функционирование нитрогеназы: кружком обозначена восстановленная форма белка
Перенос одного электрона на МоFe-белок сопровождается гидролизом как минимум двух молекул АТФ. За один раз FeS-центрами ферродоксина, Fe- и МоFe-белков может быть перенесено не более двух электронов, а для восстановления N2 до аммиака необходимо 6 электронов. Весь процесс состоит из трех последовательных стадий восстановления:
N2
![]() HN = NH
H2N
– NH2
2NH3
HN = NH
H2N
– NH2
2NH3
диимин гидрозин
Процесс азотофиксации связан с затратой большого количества энергии (для восстановления одной молекулы N2 требуется не менее 12 молекул АТФ). Помимо N2 нитрогеназа может восстанавливать ряд других субстратов, таких как N2O, C2H2 и его аналоги N3-, CN-. В отсутствие N2 нитрогеназа катализирует выделение молекулярного водорода в реакции, протекающей с затратой АТФ.
Азотофиксация обогащает азотом и почву, и растения. Органические азотсодержащие соединения в тканях растений и животных, попадая в почву, подвергаются минерализации до аммонийных соединений.
Аммонификация. Аммонификация – это отщепление аминогруппы от аминокислоты с выделением свободного аммиака в процессе дезаминирования.
Значительная часть белков поступает в почву с остатками отмерших растений, животных и микроорганизмов.
Внеклеточные протеазы микроорганизмов гидролизуют высокомолекулярные белки до пептидов различных размеров и аминокислот. В дальнейшем аминокислоты могут участвовать в синтезе белка и других полимеров, а также подвергаться переаминированию, дезаминированию или декарбоксилированию. Процесс отщепления аминогрупп с выделением аммиака, может протекать как внутри клетки, так и внеклеточно. Существует четыре типа дезаминирования:
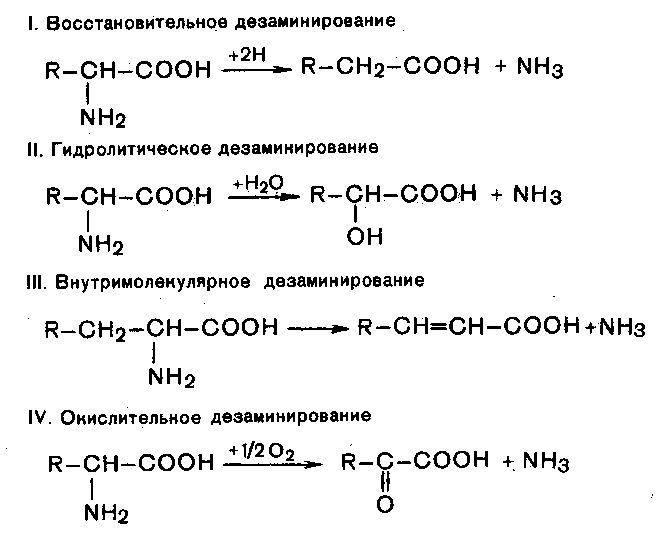
Аминокислоты минерализуются с различной скоростью. Некоторые из них (треонин, метионин) более устойчивы, другие (аргинин, триптофан), наоборот, разлагаются легко. После дезаминирования углеродный остаток подвергается воздействию микробов в аэробных и анаэробных условиях до образования СО2 и различных соединений. При аэробном распаде белка основными конечными продуктами процесса являются СО2, аммиак, сульфаты и вода. В анаэробных условиях образуются аммиак, амины, СО2, органические кислоты (жирные и ароматические – бензойная, ферулиновая и др.), меркаптаны, а также вещества с неприятным запахом – индол, скатол, сероводород.
Наиболее активные аммонификаторы – это представители грамположительных споровых бактерий из рода Bacillus. Из необразующих эндоспоры форм к аммонификаторам относятся виды грамположительных бактерий родов Micrococcus, Arthrobacter, Micobacterium, грамотрицательные виды рода Proteus, Pseudomonas, а также многие микромицеты.
Роль аммонификаторов в природе значительна, поскольку доля белка в тканях умерших растений и животных велика, а аммонифицирующие микроорганизмы осуществляют минерализацию белков, разлагая их в конечном счете до СО2, NH3 и H2S.
Аммиак, образовавшийся в процессе аммонификации, частично адсорбируется в почве, потребляется как источник азота в процессе метаболизма почвенных микроорганизмов и в аммонийной форме растениями, выделяется в атмосферу, а также окисляется в нитриты и нитраты в процессе нитрификации.
Нитрификация. В процессе нитрификации образуются окисленные формы азотистых соединений. Нитрифицирующие бактерии выделены в определителе Берджи в семейство Nitrobacteriaceae и разделены на две группы, в зависимости от процессов которые они осуществляют. Бактерии, окисляющие аммоний до нитритов (I фаза нитрификации), принадлежат к родам Nitrosomonas, Nitrosococcus, Nitrosolobus, Nitrosospira, Nitrosovibrio. Перевод нитритов в нитраты (II фаза нитрификации) осуществляют бактерии родов Nitrobacter, Nitrococcus, Nitrospira.
Процессу нитрификации предшествует поглощение NH4+ и перенос его через ЦПМ с помощью медьсодержащей транслоказы. Окисление NH4+ происходит на цитоплазматических и внутрицитоплазматических мембранах. Предполагается, что на первом этапе аммиак окисляется до гидроксиламина с помощью монооксигеназы, катализирующей присоединение к молекуле аммиака одного атома О2; второй атом О2 взаимодействует, вероятно, с НАД·Н2, что приводит к образованию Н2О:
NH3 + O2 + НАД·Н2 NH2OH + Н2 O + НАД+
Гидроксиламин далее ферментативно окисляется до нитрита:
NH2OH + О2 NO2- + Н2 O + Н+
Электроны от NH2OH поступают в дыхательную цепь на уровне цитохрома с и далее на терминальную оксидазу. Их транспорт сопровождается переносом 2-х протонов через мембрану, приводящим к созданию протонного градиента и синтезу АТФ.
Вторая фаза нитрификации сопровождается потерей 2-х электронов. Окисление нитрита до нитрата, катализируемое молибденсодержащим ферментом нитритоксидазой, локализовано на внутренней стороне ЦПМ:
NO2-
+ H2O
NO3-
+ 2H2+
+ 2![]()
![]()
Электроны поступают на цитохром а1 и через цитохром с на терминальную оксидазу аа3, где акцептируются молекулярным кислородом (рис. 45). При этом происходит перенос через мембрану 2H2+.
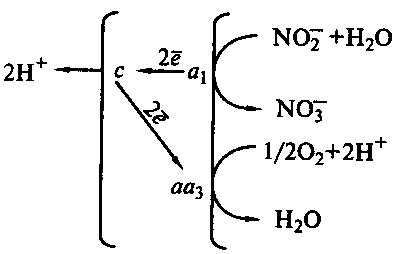
Рис. 45. Схема энергетических процессов у Nitrobacter: c, а1 , аа3 – цитохромы
Нитрифицирующие бактерии распространены в водоемах, почвах, на горных породах, на каменных и железобетонных строениях. Нитрифицирующие бактерии, как правило, развиваются совместно с микроорганизмами, осуществляющими минерализацию (аммонификацию) азотсодержащих органических веществ и приводящими к накоплению исходного субстрата нитрификации – аммиака.
Денитрификация. Денитрификация – это процесс восстановления нитратов до нитритов и далее до какой-либо из газообразных форм азота (окиси азота, закиси азота и молекулярного азота):
NO3- → NO2- → NO → N2O → N2
Способность к денитрификации обнаружена у многих почвенных и водных прокариот, среди них фото- и хемотрофы. Наиболее часто она встречается у грамположительных бактерий из родов Bacillus и Micrococcus, а также у грамположительных бактерий, принадлежащих к роду Pseudomonas. Денитрификаторы – факультативные анаэробы, переключающиеся на денитрифиакцию только в отсутствие кислорода. В аэробных условиях эти микроорганизмы осуществляют процесс кислородного дыхания.
В анаэробных условиях денитрификаторы используют нитраты и нитриты как конечные акцепторы электронов при окислении органических субстратов для получения энергии.
В процессе денитрификации каждый из четырех восстановительных этапов катализируется специфической мембраносвязывающей редуктазой. Электрон-транспортные цепи денитрификаторов в анаэробных условиях содержат флавопротеины, хиноны, цитохромы типов b и c.
Денитрификация – одна из причин обеднения почв азотом и неполного использования растениями вносимых в почву азотных удобрений. В то же время процесс имеет большое экологическое значение в сязи с тем, что он восстанавливает баланс азота, а также предохраняет водоемы от чрезмерного накопления в них нитратов, вымываемых из почв.
Денитрифицирующие бактерии широко распространены в почвах, особенно во влажных и богатых неразложившимися органическими остатками, в ризосферах, а также в илах.