

IV. 3. Изменение генетического материала
Бактерии значительно больше, чем другие организмы подвержены изменчивости. Для них характерна как наследственная, так и ненаследственная (модификационная) изменчивость. Модификационные изменения происходят на уровне фенотипа и не затрагивают клеточный генотип. Широко известны адаптивные модификации. Так, у ряда бактерий обнаружена универсальная адаптивная реакция в ответ на различные стрессовые воздействия (высокие и низкие температуры, резкий сдвиг рН и др.), проявляющаяся в интенсивном синтезе сходных белков (белков теплового шока). Адаптивные модификации расширяют возможность организма к выживанию и размножению в более широком диапазоне условий внешней среды.
В основе наследственной изменчивости лежат мутации и генетические рекомбинации.
Мутации. Почти до XX в. господствовало мнение, что в отличие от других животных бактерии выживают при неблагоприятных внешних воздействиях не благодаря случайным генетическим изменениям (мутациям), а вследствие запуска физиологических процессов, что и позволяет им выжить. Эта теория была опровергнута исследованиями С.Е. Лурия и М. Дельбрюка, которые доказали, что устойчивость E.coli к бактериальным вирусам обусловлена произошедшими в них мутациями. Исследования Лурия – Дельбрюка положили начало современной генетике микроорганизмов.
Мутации (от лат. mutare – изменять) – скачкообразные изменения в генетическом материале клетки, приводящие к появлению новых признаков.
По происхождению мутации подразделяются на спонтанные («дикие», возникающие в популяции без видимого вмешательства извне) и индуцированные (вызванные искусственно). Индуцировать мутации могут физические факторы (ультрафиолетовые и рентгеновские лучи), химические мутагены (аналоги азотистых оснований, алкилирующие и интеркалирующие агенты, азотистая кислота) и биологические факторы (транспозоны и IS-элементы). Механизм действия мутагенов различен. Выделяют мутации, также, генные (затрагивают один ген) и хромосомные (распространяются на несколько генов, связаны с крупными перестройками в отдельных фрагментах ДНК). Получение индуцированных мутаций – один из способов изучения генетики микроорганизмов.
Рекомбинация генетического материала. В 1946 г. Ледерберг и Татум (J. Lederberg, E.L. Tatum) продемонстрировали, что между членами генетически неоднородной популяции E.coli может происходить обмен генетической информацией и что при этом, как и у двуполых организмов, в результате физического обмена между хромосомами могут возникать новые генетические комбинации (генетическая рекомбинация). В настоящее время известны три способа передачи генетического материала у бактерий: конъюгация, трансформация и трансдукция.
Конъюгация – прямой перенос фрагментов ДНК от донорских клеток к реципиентам при непосредственном контакте этих клеток. Ледерберг и Татум впервые показали наличие конъюгации между бактериями. Ученые смешали два типа мутантов E.coli , один из которых нуждался в биотине и метионине, другой – в треонине и лейцине, и высеяли на минеральную среду. В результате этого появились генетические рекомбинанты со способностью к синтезу всех четырех факторов роста.
Для реализации процесса необходим F-фактор – плазмида, кодирующая информацию, необходимую для конъюгации. Конъюгация требует наличия двух типов клеток: доноров (F+), обладающих F-фактором, и реципиентов (F-), не обладающих им. F-фактор содержит гены специальных и необходимых при коньюгации структур – F-пилей и ряд других генов, вовлеченных в процесс взаимодействия с F--клетками. На первом этапе происходит прикрепление клетки-донора к клетке-реципиенту с помощью F-пилей. Затем между клетками формируется конъюгационный мостик, через который передается F-фактор. При попадании F-фактора в клетку-реципиент она становится F+ и приобретает способность предавать F-фактор другим клеткам (F+-клетки не теряют способности к передачи информации, не становятся F-клетками) (рис. 18).
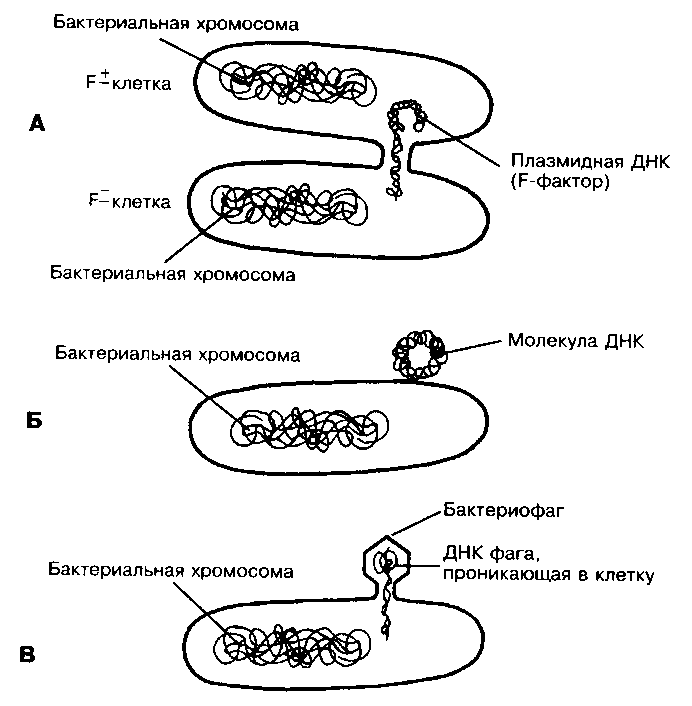
Рис. 18. Механизмы переноса бактериальной ДНК
А – конъюгация; Б – трансформация с использованием отдельной молекулы ДНК; В – трансдукция с помощью фагов
Клетки, в которых F-плазмида интегрирована в хромосому, обозначаются как Hfr+-клетками (Hfr – от англ. high frequency recombination – клетки, обеспечивающие высокую частоту рекомбинации). Hfr+-клетки способны быть донорами хромосомной ДНК. При переносе генетического материала бактериальная ДНК реплицируется, начиная от места включения F-фактора. Одна цепь ДНК переносится в реципиентную клетку F-клетку, другая остается в Hfr+-клетке, т.е. донор сохраняет свое генетическое постоянство. После начала конъюгации хромосомный материал переносится, начиная от генов, близких к начальной точке транспорта (рис. 19).
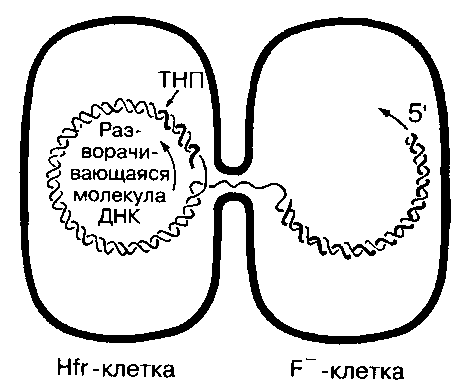
Рис. 19. Hfr-конъюгация
Hfr – бактерии функционируют при конъюгации как доноры. Перенос ДНК осуществляется линейно и начинается с удвоения места включения F-фактора (то есть с точки начала переноса – ТНП)
В бактерии-реципиенты обычно попадают первые из переносимых генов, размер которых зависит от времени, в течение которого происходила конъюгация, последними переходят гены, кодирующие F-пили. Конъюгация может быть прервана обычным встряхиванием суспензии бактерий, поэтому полный перенос генов – редкое явление и реципиентная клетка остается F-. Вслед за процессом переноса в клетке-реципиенте происходит гомологичная рекомбинация между донорской ДНК и собственной ДНК реципиента.
Трансформация – генетическое изменение клеток в результате включения в их геном экзогенной ДНК (рис. 18, Б). Погибшие клетки постоянно высвобождают ДНК, которая может быть воспринята бактериями (как правило, любая чужеродная ДНК, попадающая в бактериальную клетку, расщепляется эндонуклеазами, но при некоторых условиях такая ДНК может быть включена в геном бактерии).
Трансдукция – перенос генов от одной бактериальной клетки к другой посредством бактериофага (рис. 18, В). Трансдуцирующий бактериофаг обычно переносит небольшой фрагмент ДНК хозяина от клетки-донора к клетке-реципиент. Известно три вида трандукции: неспецифическая, специфическая и абортивная.
При неспецифической трансдукции фагом может быть перенесен любой фрагмент ДНК хозяина. Трансдуцирующие фаговые частицы образуются в ходе определенных инфекционных процессов, когда ДНК клетки деградирует и фрагменты, по размеру примерно соответствующие фаговому геному, случайно упаковываются в зрелые частицы бактериофага. В результате последующего инфицирования клеток бактерий популяций фаговых частиц, содержащих трандуцирующие фаги, происходит передача ДНК донорских клеток этим инфицируемым клеткам. Рекомбинация между введенными фрагментами донорской ДНК и ДНК клетки-реципиента приводит к изменению генотипа последней.
Для специфической трансдукции характерен перенос определенных фрагментов. С помощью такого механизма трансдуцируются только те гены клетки-хозяина, которые тесно сцеплены с сайтом интеграции вирусного генома.
Примером специфической трансдукции служит передача с помощью бактериофага . Этот фаг при переходе в состояние профага включается в геном бактерий в специфический хромосомный локус. При вырезании фаг может включать хромосомные гены, прилегавшие к интегрированному вирусному геному. Так, фаги трансдуцируют гены, ответственные за метаболизм галактозы ( gal), или гены, контролирующие синтез биотина ( bio), а фаги ф80 – различное число генов, кодирующие ферменты биосинтеза триптофана.
При абортивной трансдукции внесенный фрагмент ДНК донора не встраивается в хромосому клетки-реципиента, а остается в цитоплазме, транскрибируется, но не реплицируется. При клеточном делении передается только одной из дочерних клеток (наследуется однолинейно) и затем теряется в потомстве.