

IV. 2. Репликация днк прокариот
Для сохранения уникальных свойств организма необходимо точное воспроизведение генетической информации в каждом последующем поколении. Во время деления клетки содержание ДНК должно удвоиться (реплицироваться), чтобы каждая дочерняя клетка могла получить полный спектр ДНК. В основе репликации лежит матричный механизм биосинтеза. Во время репликации каждая из цепей ДНК служит матрицей для синтеза комплементарной дочерней цепи. После репликации одна цепь в каждой из двух дочерних молекулах ДНК является родительской, т.е. консервативной, а другая синтезируется заново. Такой способ удвоения молекул ДНК называется полуконсервативным.
Репликация лучше изучена у кишечной палочки. Хромосома E.coli имеет кольцевую форму и содержит около четырех миллионов пар оснований. В репликации выделяют три стадии: инициацию (начало синтеза), элонгацию (продолжение), терминацию (окончание синтеза).
Инициация репликации. В определенной точке (точке начала репликации – точка ori), цепи расходятся и образуют две репликативные вилки, которые движутся в противоположных направлениях до тех пор, пока не встретятся на противоположной стороне кольца (рис. 16). ДНК синтезируется со скоростью 500 пар оснований в секунду.
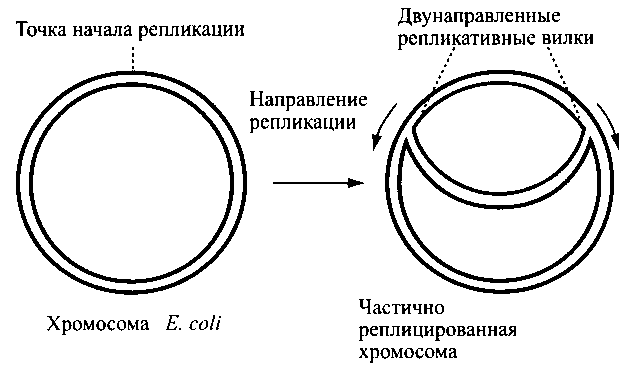
Рис. 16. Двунаправленная репликация хромосомы E.coli
Модель полуконсервативной прерывистой репликации ДНК в репликативой вилке представлена на рис. 17. Дуплекс родительской молекулы ДНК расплетает фермент АТФ – зависимая хеликаза. Образующиеся одноцепочечные участки кооперативно покрывает SSB-белок (от англ. Single Strand Binding Proteins). SSB-белки стабилизируют одноцепочечные ДНК, защищают от обратимого отжига. Когда нити используются для синтеза новой ДНК, SSB-белки удаляются.
Синтез новой цепи ДНК невозможен без затравки, или праймера (от англ. primer – затравка), короткого олигодезоксирибонуклеотида или олигорибонуклеотида (до пяти нуклеотидов), комплементарного соответствующему участку ДНК-матрицы, у которого имеется свободная 3/-ОН-группа. Затем короткие праймеры замещаются сегментами ДНК.
Элонгация репликации. После синтеза праймера присоединяется ДНК-полимераза III , которая начинает синтез новой цепи. ДНК-полимеразы способны наращивать цепь только в направлении 5/ 3/, поэтому только на одной цепи, ориентированной в направлении 3/ 5/, синтез цепи может проходить непрерывно (ведущая, лидирующая цепь). На второй матричной цепи, ориентированной в направлении 5/ 3/, синтез ДНК синтезируется короткими фрагментами (1000–2000 нуклеотидов), называемыми фрагментами Оказаки, в честь открывшего их ученого. Цепь, собираемая из фрагментов, называется ведомой, или отстающей. Синтез каждой из фрагментов отстающей цепи начинается с затравки, которые через некоторое время удаляются, бреши застраиваются ДНК-полимеразой I и фрагменты сшиваются специальным ферментом ДНК-лигазой в непрерывную цепь.
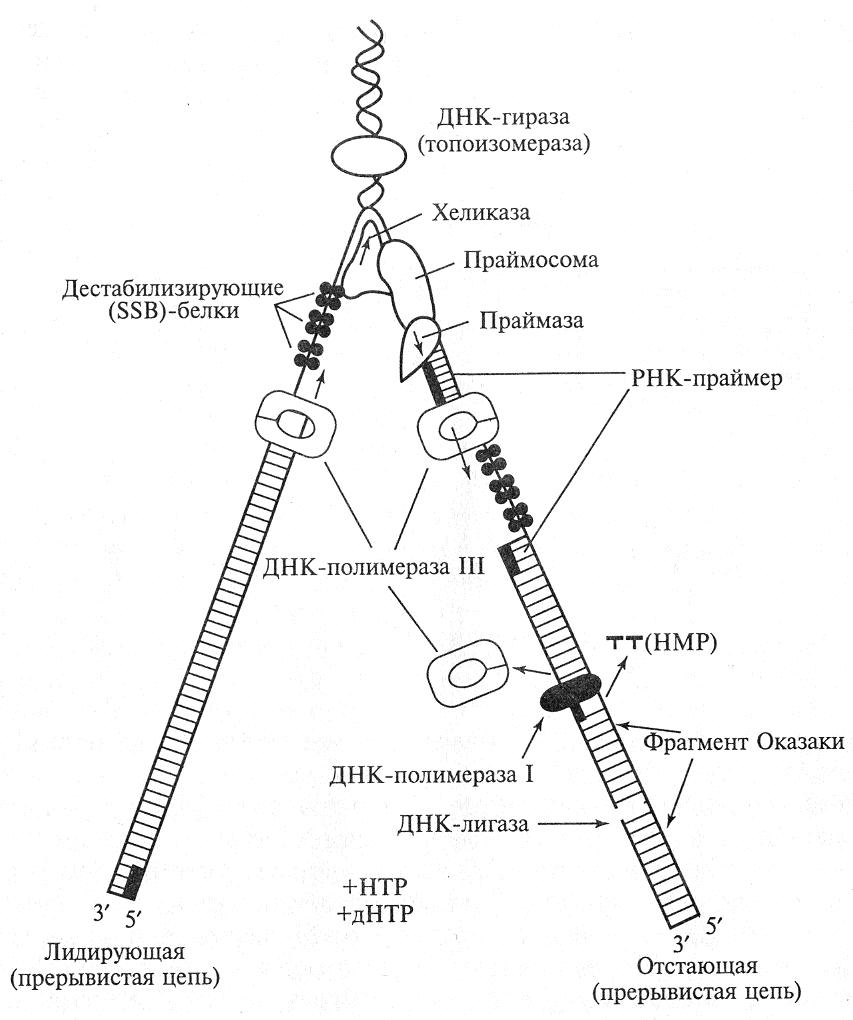
Рис. 17. Схема синтеза ДНК в репликативной вилке у прокариот
Терминация репликации. Окончание репликации происходит тогда, когда встречаются две репликативные вилки при удвоении кольцевых молекул ДНК. Кольца в этих местах встречи соединяются ДНК-лигазой, при этом обычно они оказываются попарно сцепленными, т.е. образуют катенат. ДНК-гираза вносит временный двуцепочечный разрыв и расцепляет зацепленные кольца.