

Просторове розташування субодиниць рибосоми під час трансляції.
Генетичний код – система запису інформації про амінокислоти за допомогою триплетів нуклеотидів в молекулі ДНК або РНК. Генетичний код триплетний. Кожна амінокислота зашифрована кодоном – послідовністю із трьох нуклеотидів. Загальна кількість кодонів – 64 (43). Важливими для кодування є два перші нуклеотиди в кодоні, тоді як роль третього не є суттєвою. Генетичний код є виродженим – кожна амінокислота шифрується більш ніж одним кодоном (від 2 до 6). Генетичний код є однозначним, тобто кожний кодон кодує тільки одну амінокислоту. Генетичний код є універсальним – він є загальним для всіх організмів. Проте із цього правила є виключення. Так, код білкового синтезу в рибосомах мітохондрій відрізняється від системи кодування у всіх інших рибосомах прокаріот та еукаріот.
Функціональні характеристики гену:
гени є ділянками ДНК,
один ген кодує синтез одного білка (моногенні ознаки),
гени взаємодіють один з одним (полігенні ознаки).
дія гену специфічна – кодує одну амінокислотну послідовність,
деякі гени мають властивість плейотропність,
дозованість дії гену залежить від інтенсивності прояву ознаки та кількості певного алеля,
на активність гену впливають як зовнішнє, так і внутрішнє середовище,
конститутивні гени постійно експресуються, неконститутивні гени неактивні.
Експресія генів – реалізація генетичної інформації (ДНК → про-іРНК → іРНК → білок → модифікації білкової молекули).
Регуляцію експресії генів розглянемо на прикладі лактозного оперону у Escherichia coli. Подібний механізм регуляції є у більшості прокаріотів; у еукаріотичних організмів цей механізм є значно складнішим.
Цитоплазматична спадковість - позаядерна спадковість, яка забезпечується молекулами ДНК, які містяться в пластидах і мітохондріях. Генетичний вплив цитоплазми є наслідком взаємодії плазмону з ядерними генами. Ознака, що визначається цитоплазмою, успадковується по материнській лінії.
Плазмон – сукупність генів мітохондрій.
Пластом – сукупність генів пластид.
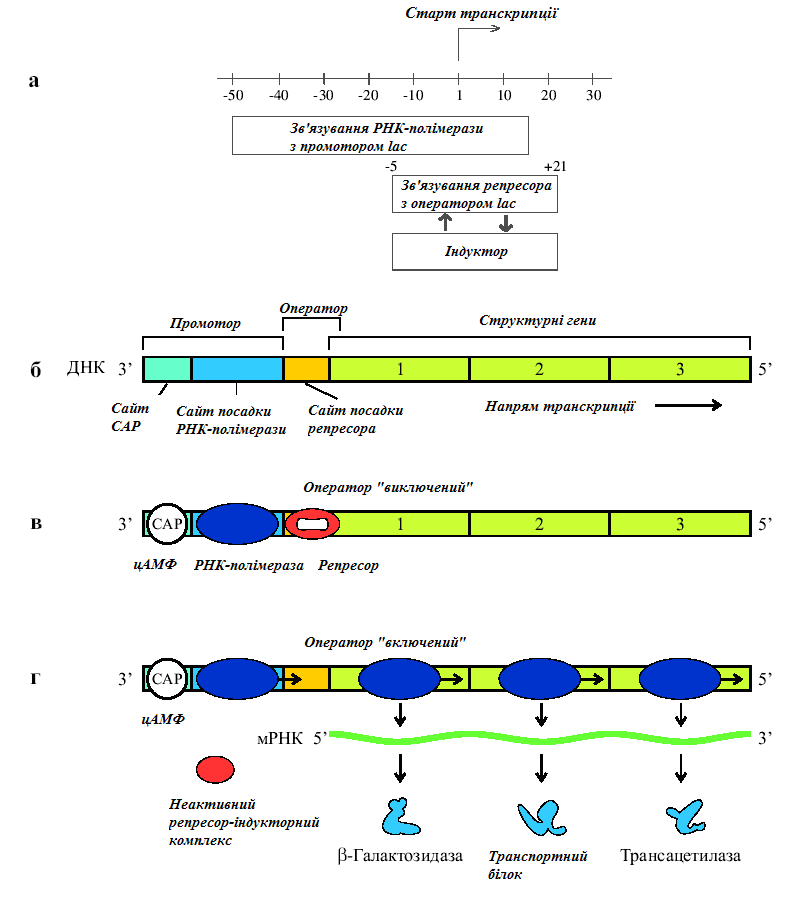
Функціонування лактозного оперону у e.Coli.
а. Локалізація сайтів зв’язування молекул РНК-полімерази і репресора в регуляторній області гену lacZ (Із: Lewin, 1994, p.417). б. Структура лактозного оперону. Як і у всіх генів, що регулюються САР і сАМР, промотор містить два райони: ділянку зв’язування з РНК-полімеразою і ділянку зв’язування з комплексом САР-сАМР. в, г. Негативна і позитивна регуляція lac-оперону. За відсутності лактози репресор (продукт гена І) зв’язується з оператором. Хоча РНК-полімераза може зв’язуватися з промотором, вона не може переміщуватися далі репресора. Оператор виключений, гени не працюють. За присутності індукуючої молекули репресор інактивується і більше не зв’язується з оператором. Молекули РНК-полімерази переміщуються, і починається транскрипція (б-г – із: Curtis, Barnes, 1989, p.325).
Тема 4. Генетична мінливість. Мутації. Генетична інженерія.
Основні типи мінливості:
1. спадкова – обумовлюється змінами, які відбуваються в структурах матеріальних носіїв спадковості – генах або хромосомах. Зміни успадковуються, характеризують мутантні властивості фенотипового прояву.
2. неспадкова (модифікаційна) – обумовлюється реакцією генотипів організмів на умови зовнішнього середовища.
3. комбінаційна – виникає внаслідок пере комбінації неалельних генів. Є результатом незалежного вільного розподілу хромосом в процесі редукційного поділу клітини, кросинговеру, який змінює співвідношення між домінантними і рецесивними неалельними генами в межах кожної хромосоми, а також внаслідок генетичної трансформації, трансдукції, сексдукції.
Норма реакції – це зміна ознак чи властивостей організму, що виникає під впливом умов зовнішнього середовища і не передається нащадкам. Механізм виникнення модифікацій – геномодуляція (зміна функцій генів).
Основні тези мутаційної теорії Г. де Фріза:
1. мутація виникає раптово, без всяких перехідних форм;
2. нові форми, що виникли внаслідок мутацій, є досить стійкі;
3. в протилежність не спадковим змінам мутації не утворюють безперервних рядів, не групуються навкруги середнього типу;
4. мутації проявляються по-різному і можуть бути як корисними, так і шкідливими;
5. вірогідність виявлення мутацій залежить від кількості досліджених особин;
6. одні і ті ж мутації можуть виникати багаторазово.
Основні відмінності мутацій і модифікацій.
№ |
Особливості мутацій |
Особливості модифікацій |
1. |
Невизначеність |
Визначеність |
2. |
Вираженість змін не залежить від сили і тривалості дії фактора, що викликає мутації |
Ступінь змін фенотипу прямо пропорційний силі і тривалості впливу провокуючого фактора |
3. |
Не мають безпосереднього адаптаційного значення. Інколи можуть бути корисними, але лише випадково |
У переважній більшості мають адаптивне значення в межах норми реакції. Виключенням з цього правила є переважна більшість морфозів. |
4. |
Константні (не зникають протягом життя особин) |
Не стійкі. Як правило, зникають протягом життя особини. Виключення – тривалі модифікації. |
5. |
Успадковуються |
Не успадковуються |
Класифікація мутацій.
1. Залежно від способу виникнення мутацій:
а/ спонтанні, що постійно виникають у природі без очевидних причин і з певною частотою;
б/ індуковані мутації, що виникають у відповідь на дію різноманітних факторів середовища.
2. За виявом у гетерозиготи:
а/ домінантні мутації;
б/ рецесивні мутації.
3. За відношенням до норми або так званого дикого типу:
а/ прямі мутації, за яких гени дикого типу перетворюються в алель ні форми;
б/ суп ресорні і зворотні мутації, за яких відновлюється дикий фенотип.
4. За локалізації в еукаріотній клітині:
а/ ядерні, якщо мутації відбуваються в ДНК ядра;
б/ цитоплазматичні, якщо мутації відбуваються в ДНК цитоплазми.
5. В залежності від типу клітин, в яких виникають мутації:
а/ генеративні – виникають у статевих клітинах та їх попередниках;
б/ соматичні – виникають у соматичних клітинах і розповсюджуються за їх мітотичного поділу.
6. За фенотиповим виявом:
а/ морфологічні – мутації, що проявляються тими чи іншими змінами будови клітин та організмів, структури колоній прокаріотів тощо;
б/ фізіологічні – супроводжуються порушенням фізіологічних функцій;
в/ біохімічні – мутації, для яких встановлена суть основних порушень обміну речовин, в першу чергу на рівні білкових молекул.
7. За впливом на адаптивну здатність клітин і організмів:
а/ корисні мутації – за фенотиповим проявом імітують адаптивні модифікації і тому сприяють збереженню виду за даних умов;
б/ нейтральні мутації – не впливають на життєздатність клітин і організмів;
в/ субвітальні мутації – знижують життєвість генотипів на 10-50%;
г/ напівлегальні мутації – знижують життєвість генотипів на 50-90%;
д/ летальні мутації – призводять до загибелі 100% генотипів, що мають таку мутацію;
є/ умовно-летальні мутації – проявляються лише за певних умов.
8. Залежно від змін генотипу:
а/ генні або точкові мутації – зміни структури ДНК у межах гена.
Мутація, що виникає в разі заміни в молекулі ДНК пуринової основи на пуринову, або піримідинової на піримідинову, називається транзицією (А↔Г, Т↔Ц). Мутація внаслідок заміни пуринової азотистої основи на піримідинову, і навпаки, називається трансверсією (А↔Т, Г↔Ц, А↔Ц, Г↔Т). До точкових мутацій також відносять вставки (інсерції) одного або декількох зайвих нуклеотидів, випадіння (делеції) одного або декількох нуклеотидів, перестановки сусідніх нуклеотидів. Заміни одного нуклеотида на інший в триплеті можна поділити на нейтральні (смислове значення кодону не змінюється в зв’язку з виродженістю генетичного коду), місенс- мутації (призводять до зміни змісту відповідного триплету (кодону)) та нонсенс-мутації (спричиняють припинення процесу біосинтезу білка).
б/ хромосомні мутації (хромосомні перебудови, аберації) – порушення структури хромосом.
Х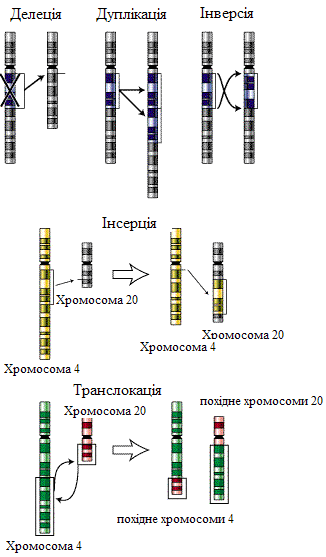 ромосомні
мутації поділяються на внутрішньохромосомні
та міжхромосомні:
1) делеції
або
нестачі
певної ділянки хромосоми, 2) дуплікації
(одна із ділянок хромосоми представлена
у вигляді двох або значно більшої
кількості копій); 3) інверсії
(в одній із ділянок хромосоми гени
розташовані в зворотній послідовності);
4) інсерції
(фрагменти хромосомної нитки з одного
місця переносяться в інше місце тієї ж
або іншої хромосоми, зберігаючи вихідний
порядок розміщення генів в межах
фрагменту); різновидом інсерцій є
транспозиції;
5) транслокації
(реципроктні (взаємні) обміни між
негомологічними хромосомами).
ромосомні
мутації поділяються на внутрішньохромосомні
та міжхромосомні:
1) делеції
або
нестачі
певної ділянки хромосоми, 2) дуплікації
(одна із ділянок хромосоми представлена
у вигляді двох або значно більшої
кількості копій); 3) інверсії
(в одній із ділянок хромосоми гени
розташовані в зворотній послідовності);
4) інсерції
(фрагменти хромосомної нитки з одного
місця переносяться в інше місце тієї ж
або іншої хромосоми, зберігаючи вихідний
порядок розміщення генів в межах
фрагменту); різновидом інсерцій є
транспозиції;
5) транслокації
(реципроктні (взаємні) обміни між
негомологічними хромосомами).
в/ геномні мутації – випадкові зміни кількості окремих хромосом або кількості хромосомних наборів. До таких мутацій відносять: 1) центричне злиття (дві негомологічні хромосоми зливаються в одну); 2) центричний поділ (одна хромосома поділяється на дві, при цьому обов’язково виникає одна нова центромера); 3) аутополіплоїдія (збільшення в клітині кількості хромосом, кратне гаплоїдному наборові); 4) алополіплоїдія (у випадку, коли між собою спонтанно схрещуються особини, які належать до різних таксономічних видів спорідненого походження); 5) анеуплоїдія (в хромосомному наборі відсутня одна чи декілька хромосом, або ж, навпаки, з’являється одна або декілька додаткових).
Особливості мутацій цитоплазматичних генів: 1) успадковуються по материнській лінії; 2) велика кількість екземплярів; 3) здатність (або нездатність) нової мутації створювати з сусідніми генами життєздатну комбінацію. Приклади: мутації стійкості клітин прокаріотів до антибіотиків, хлорофільні мутації, мутації інтенсивності газообміну (дихання), мутації темпів росту, мутації чоловічної цитоплазматичної стерильності, мутації статевої несумісності.
Репарація – особлива функція клітин, яка полягає у здатності виправляти хімічні пошкодження і розриви в молекулах ДНК, що пошкоджена при нормальному біосинтезі ДНК в клітині або в результаті впливу фізичних або хімічних агентів. Здійснюється спеціальними ферментними системами клітини. З порушеннями систем репарації пов’язаний цілий ряд спадкових хвороб (наприклад, пігментна ксеродерма). Кожна із систем репарації включає наступні компоненти: 1) фермент, що “впізнає” хімічно змінені ділянки в ланцюзі ДНК і здійснює розрив ланцюга поблизу від місця пошкодження; 2) фермент, що видаляє пошкоджену ділянку; 3) фермент (ДНК-полімераза), який синтезує відповідну ділянку ланцюга ДНК замість видаленої; 4) фермент (ДНК-лігаза), який замикає останній зв’язок в полімерному ланцюзі і тим самим відновлюючи його безперервність.
Закон гомологічних рядів у спадковій мінливості організмів (М.І.Вавілов, 1920 р.):
1. Види і роди, генетично близькі, характеризуються подібними рядами спадкової мінливості з такою правильністю, що, знаючи ряд форм у межах одного виду, можна передбачити існування паралельних форм у інших видів і родів. Чим ближче генетично розташовані в загальній системі роди і види, тим повніша подібність у рядах їхньої мінливості.
2. Цілі родини рослин взагалі характеризуються певним циклом мінливості, яка проходить через усі роди й види, що утворюють родину.