

1.3 Инфицирование и развитие микозов
Проникновение гриба в организм насекомых-хозяев может происходить практически через любые участки тела: кишечник, трахеи, поровые канальцы в кутикуле, склериты, межсегментарные участки, но основной путь попадания – через кутикулу насекомого[1].
Кутикула выполняет функцию экзоскелета, защищает организм насекомого от иссушения и служит барьером между внешней средой и организмом. Она покрывает все отделы насекомого, за исключением среднего кишечника. Кутикула состоит из двух основных частей: верхней – эпикутикулы и нижней прокутикулы в соответствии с рисунком 4. Эпикутикула, толщиной примерно (0,1 – 3) мкм, определяет микрорельеф покровов и селективную проницаемость для различных веществ. Самый внутренний слой её состоит из белков и липидов и носит название «гомогенная протеиновая эпикутикула». Протеиновый слой первым образуется при формировании новой кутикулы. Белки верхней части, задубленные хинонами, образуют кутикулин.
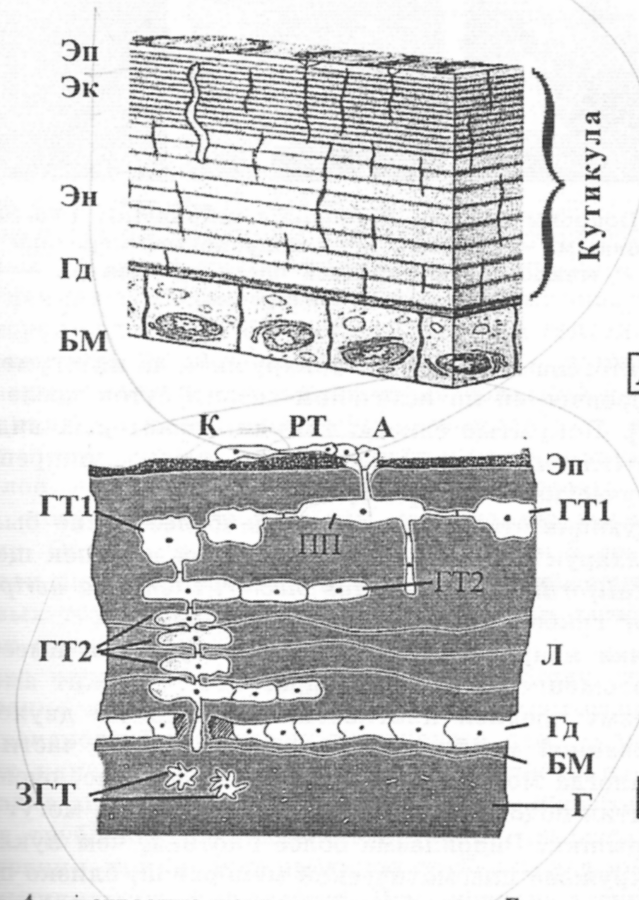
Эп – эпикутикула, Эк – экзокутикула, Эн – эндокутикула, Гд – гиподерма, БМ – базальная мембрана, К – конидия, РТ – ростовая трубка, А – апрессорий, ПП – проникающая плата, ГТ1 – гифальные тела первого порядка, ГТ2 – гифальные тела второго порядка, ЗГТ – звездообразные гифальные тела, Л – ламеллы, Г – гемолимфа
Рисунок 4 – Строение кутикулы и этапы инфицирования насекомых (сверху вниз)
Протеиновый слой и кутикулин легкопроницаемы для воды и низкомолекулярных неполярных соединений. Защита организма от высыхания обеспечивается восками, покрывающимим белковый слой эпикутикулы. Под восками понимают смесь предельных и непредельных углеводородов, продуктов их окисления: спиртов, жирных кислот, эфиров жирных кислот, стероидов. Воска синтезируются эпидермальными клетками. Их состав зависит от вида насекомого и условий окружающей среды. Толщина прокутикулы может варьировать от десяти до нескольких сотен микрон. Она обуславливает механические свойства кутикулы, то есть фактически выполняет роль экзоскелета. Её основу составляют хитин и белки, на долю которых приходится около 90% сухой массы. Первый построен из мономеров N-ацетилглюкозамина, и синтезируется гиподермой и клетками гемолимфы из глюкозы. В кутикуле хитин за счет водородных связей между соседними полимерными цепями образует микрофибриллы, которые объединяясь, дают начало более крупным структурам – нитям или пучкам. Микрофибриллы способны адсорбировать водорастворимые кутикулярные белки. Хитин, вступая в связь с белком, сильно изменяет свои свойства – приобретает устойчивость к действию хитиназ, его микрофибриллы становятся менее эластичными. Хитин-белковый комплекс образует чередующиеся слои, состоящие из тонких пластинок – ламелл. Прочность кутикулы значительно возрастает после её склеротизации – процесса образования тесной связи между белками и хинонами. В затвердевшей кутикуле такие белки теряют способность к растяжению, превращаясь в прочные склериты. Процессы склеротизации в основном идут в верхней части прокутикулы, поэтому она часто именуется экзокутикулой, тогда как лежащая под ней часть – эндокутикулой в соответствии с рисунком 4.
Далее за кутикулой располагается эпидермис или гиподерма, которая также является мишенью гриба-патогена на более поздних стадиях заражения. Она состоит из одного слоя вытянутых или многоядерных клеток. В гиподерме локализованы клетки, дающие начало волоскам или сенсиллам, а также различные железы. Многие эпидермальные клетки содержат гранулы пигментов. Другие функции – синтез различных биологически-активных веществ: пигментов, строительного материала для кутикулы и т.д. Базальная мембрана – аморфный гранулярный слой, отделяющий эпидермис от полости тела, не имеет клеточного строения. Она секретируется клетками гемолимфы и состоит из мукополисахаридов.
Инфицирование насекомых происходит в несколько этапов в соответствии с рисунком 4:
1 адгезия;
2 развитие гриба на поверхности кутикулы;
3 проникновение и развитие гриба в кутикуле;
4 вторжение в полость тела и колонизация внутренних органов насекомого[1].
Адгезия конидий грибов на кутикуле происходит за счет гидрофобных взаимодействий между липидными компонентами эпикутикулы и клеточной оболочки, а также за счет неровностей и выростов (щетинки, складки и др.) на поверхности кутикулы. Последний фактор играет значительную роль, поскольку в момент прорастания споры обнаруживаются в основном в различных складках, где могут сохраняться длительное время.
Типичная непроросшая конидия имеет круглое, расположенное в центре ядро. В начале или незадолго до прорастания ядро становится вытянутым и перемещается из центра споры к полюсу прорастания, где начинает делиться. Через некоторое время у одного из полюсов появляется ростовая трубка. Одно из дочерних ядер после деления мигрирует в ростовую трубку. Оставшееся в конидии ядро со временем разрушается. Большинство клеточных органелл перемещается в ростовую трубку. Прорастание считается законченным, когда ростовая трубка отделяется септой от споры. В месте соприкосновения с покровами ростовая трубка начинает разрастаться, образуя аппрессориальную клетку, которая отделяется от ростовой трубки септой. Закончив свое формирование, аппрессориальная клетка начинает делиться. Дочерние клетки отделяются от материнской септами. Аппрессориальные клетки покрыты слизеподобным веществом, которое и обеспечивает прочное сцепление этих структур с эпикутикулой.
Перед началом внедрения гриба в кутикулу наблюдается образование полостей в эпикутикуле под аппрессорием. При этом происходит исчезновение воскового слоя и расширение поровых канальцев. В образовавшиеся полости переливается цитоплазма аппресориальных клеток, затем образуется проникающая трубочка. При этом вокруг проникающей трубочки деформация слоев эпикутикулы не наблюдается. Исчезновение воскового слоя, расширение поровых канальцев и отсутствие видимого механического повреждения эпикутикулы обусловлено действием гидролитических ферментов гриба в процессе инфицирования. Поскольку главными компонентами кутикулы насекомых являются липиды, белки и хитин, то основными ферментами, разрушающими кутикулу, являются липазы, протеазы и хитиназы. Проникающая трубочка, достигнув прокутикулы, начинает разрастаться параллельно её ламеллам, в результате чего формируется дисковидная структура – проникающая плата. Митохондрии, эндоплазматический ретикулум, липоидные гранулы, рибосомы, диктиосомы и ядерный аппарат клетки в проникающую плату переходят из проникающей трубочки. После миграции органелл на уровне эпикутикулы появляется септа. После этого из проникающей платы начинают развиваться отростки, растущие вниз и латерально. Эти отростки дают начало гифальным телам первого и второго порядка. Гифальные тела имеют неправильную форму, плотную цитоплазму и тонкую клеточную стенку. Гифальные тела второго порядка – правильной формы, овальной или вытянутой; цитоплазма менее плотная, клеточная стенка более толстая. Образование гифальных тел происходит путем почкования или гифальных тел, или латеральных гиф. Первоначально развитие гриба происходит в верхней части прокутикулы. Гифальные тела второго порядка продуцируют короткую вертикальную гифу, достигающую пространства между двумя смежными ламеллами и дающую начало новым гифальным телам второго порядка. В нижележащих слоях прокутикулы вертикальная гифа имеет тенденцию к удлинению и продуцированию нескольких гифальных тел.
В местах внедрения гриба в гиподерму наблюдается лизис клеток этой ткани. Гифы, проникшие за пределы этой ткани, увеличиваются в размере и начинают делиться. Начальные этапы колонизации внутренних органов характеризуются формированием в гемолимфе звездообразных гифальных тел. Рост и размножение гриба не очень интенсивны, пока хозяин жив; большинство гифальных тел концентрировалось в местах проникновения. Интенсивная колонизация внутренних органов насекомого наблюдается после его гибели. В первую очередь гриб атакует жировое тело, затем наступает очередь кишечного тракта и мальпигиевых сосудов. Остальные органы заселяются в следующем порядке: гиподерма – нервная система – мышечная ткань – трахеи. Через некоторое время гифы начинают расти в обратном направлении – к поверхности тела. Следует отметить, что очевидного разрушения органов и тканей, и особенно прокутикулы, во время роста гриба к поверхности тела может не происходить. Выход гиф на поверхность осуществляется путем разрыва покровов гифами, находящимися в прокутикуле. Споруляция начинается через сутки после появления вегетативных гиф.
Развитие энтомопатогенных грибов в организме сопровождается гистолизом тканей и органов вокруг проникающих грибных элементов. Активную роль в этом процессе играют ферменты патогена. Уникальная способность грибов проникать в организм насекомых через кутикулу в значительной степени обусловлена наличием ферментов, разрушающих кутикулярный материал [1]. Поскольку главными компонентами кутикулы являются липиды, белки и хитин, то основными ферментами патогена являются липазы, протеазы и хитиназы. Было изучено, что гриб Verticillium lecanii обладает протеазной, аминопептидазной, карбоксипептидазной, липазной, эстеразной, хитиназной, N-ацетилглюкозаминазной, каталазной, дезоксирибонуклеазной, рибонуклеазной активностью. Во время развития гриба гидролизу сперва подвергается белковая часть кутикулы, и только потом начинается разрушение хитина.
Смерть насекомого происходит до интенсивной колонизации его органов и тканей. Обусловлено это с одной стороны ферментами, а с другой – продуцированием токсинов грибом, а также токсическими продуктами распада тканей хозяина.
Энтомопатогенный гриб V. lecanii синтезирует широкий спектр метаболитов: соединения дипсипептидной природы (афидиколин, боверицин, бассианолид), 2,6-диметоксибензохинон и органические кислоты (С25 – соединения, производные дипиколиновой кислоты и пр.) активные перорально и при инъекциях [4]. Боверицин – это циклодепсипептид, состоящий из четырех молекул, каждая из которых включает N-метиллизин и D-α-гидроксиизовалериановую кислоту [5]. Из мицелия гриба Verticillium lecanii выделены и частично охарактеризованы инсектицидные метаболиты липидной природы, проявляющие контактную активность в отношении сосущих насекомых [6].