Клонирование генов эукариот.
Прокариоты не могут удалять интроны(некодирующиепосл-ти) из первичных РНК-транскриптов => верная трансляция (синтез белка на матрице РНК) эукар. мРНК в бакт.клетке невозможна. Экспрессия эукар. ДНК осущ-сятолько при наличии прокар. сигнальных посл-тей, регулир. транскрипцию и трансляцию. Концевые участки эукар.мРНКмодифиц-ны: их 5'-концы кэпированы (присоединены концевые посл-ти из остатка гуанина, часто метилированного), а 3'-концы полиаденилированы («хвост» из 200 остатков аденозина). Наличие poly(А)-хвоста позволяет отделить мРНК от р- и т-РНК. Для этого всю РНК клетки пропускают через колонку сцеллюлозой, к кот.«пришиты» oligo(dT) - короткие олигонуклеотидные цепочки из остатков тимидина(около 15 звеньев).Poly(A)- хвосты мРНК спариваются с oligo(dT) и задерж-ся в колонке, атРНК и рРНК проходят через нее. Затем колонку промывают буфером=> разрыв водородных связей между аденином и тимином, и мРНК высвобождается.
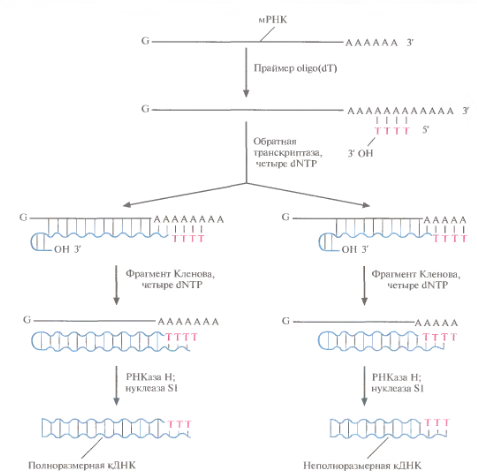
Саму мРНК нельзя встроить в ДНК-вектор, сначала на ней необх. синтез. ДНК. Для этого исп-ют 2 полимеразы: обратную транскриптазу (ОбрТр) и фрагмент Кленова ДНК-полимеразы I (рис.). Вначале в реакц. смесь с очищенной мРНК добавляют короткиеoligo(dT), ОбрТри 4 типа дезоксирибонуклеотид-3-фосфата. Ро1у(А)-хвостмРНК спаривается с oligo(dT), несущим свободную 3'-ОН-группу, кот. инициирует синтез комплементарной цепи. Матрица – мол-ла мРНК, а катализирует егоОбрТр, котораяпослед-но присоединяет к растущей цепи остатки Т, С, G или А, комплем.нуклеотидам мРНК. Invitro синтез ДНК идет не до конца,фермент перед остановкой «поворачивает назад» и присоед. несколько нуклеотидов в обратном направлении, образуя «шпильку».
В смесь добавляют фрагмент Кленова ДНК-полимеразы I Е. coli, который достраивает 2ю цепь ДНК, исп-я 1ю как матрицу. Он присоед. дезоксирибонуклеотиды к растущей цепи, начиная с З'-ОН-конца шпильки. По окончании синтеза препарат обраб-ют ферментом РНКазой Н, разрушающеймРНК, и нуклеазой S1, отщепляющей 1-цепочечные концы ДНК. Получ. препарат - смесь частично и полностью 2-цепочечных комплем. ДНК-копий (кДНК) мРНК исходного образца.
РазныекДНК можно встроить в плазмидный вектор и получить кДНК-библиотеку. Для скрининга кДНК-библ. (отбора клонов с гибриднымиплазмидами), исп-ют метод гибридизации или иммунологические методы. В последнем кДНКд.б встроена в сайт, нах-ся под контролем бактер. промотора, обесп. транскрипцию.
Рис. Синтез кДНК. К препарату очищенной мРНК добавляют праймерoligo(dT). Для синтеза ДНК на РНК-матрице исп-ют фермент обратную транскриптазу и четыре dNTP. Invitro обратная транскриптаза не обеспеч. синтез полноразмерных кДНК-копий на всех матрицах и образ. на конце растущей цепи шпильку с З'-ОН-группой. Эта группа инициирует синтез 2й цепи ДНК при участии фрагмента Кленова. После завершения синтеза молекулы мРНКгидролизуютРНКазой Н, а ДНК обраб. нуклеазой S1, получая линейные молекулы ДНК с тупыми концами без шпилек.
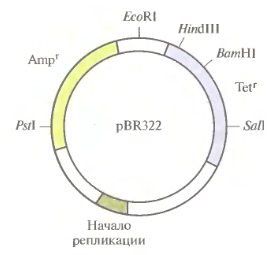
Плазмидный вектор pBr 322.
pBR322-вектор, особ. популярный в 80е гг. Р-(от англ.plasmid),BR-авторы Боливар и Родригес,322-№ записи в протоколе. Длина-4361 п. н. Состав вектора: - 2 гена резистентности к а/б ампициллину и тетрациклину (Аmрг и Tetr – сайты рестрикции в области генов устойчивости к этим а/б); - 5 уникальных сайтовузнавания длярестриктаз (ферментов, разрезающих ДНК): ВатНI, HindIII иSalIв гене Теtr,PstI-сайт в гене Аmрг,EcoRI – вне кодирующих после-тей; - сигнал начала репликацииorigin. Репликацияосущ-сятолько в Е. coliс образ.мн-вакопий, в другие бактерии переносится с трудом.
Вектор популярен, т.к. гены резистентности к а/б – распростр. селективные маркеры.Aмпициллин убивает растущие бактер. клетки, блокируя заключит. стадию синтеза клет. стенки; ген устойч. к Ampкодирует ферм. β-лактулозу, кот. расщепл. β-лактамовое кольцо а/б. Teтрациклин блокирует синтез белка в бактер.клетке, связ. С 30S-субъед. рибосом; ген Tetrкодирует белок, модиф-щий мембраны бактерий, препятствуя проникн. а/б в клетку. Клетки с генами устойч. к Amp и Tet в составе р-ДНК (вектор) приобретают спос-ть расти на среде с а/б, хотя реципиентные клетки д.б. чувствительны к этим а/б, т.е. AmpS(sensitive).