

Электрофизиологические характеристики миокардиальных клеток
Проводящие и рабочие кардиомиоциты являются возбудимыми структурами, т.е. обладают способностью к генерации и проведению потенциалов действия (нервных импульсов). Причем для проводящих кардиомиоцитов свойственна автоматия (способность к самостоятельной периодической ритмической генерации нервных импульсов), тогда как рабочие кардиомиоциты возбуждаются в ответ на приходящее к ним возбуждение от проводящих или других уже возбужденных рабочих миокардиальных клеток.
Первоначально в закладывающемся сердце эмбриона все кардиомиоциты обладают автоматией, для них характерно слабое развитие миофибриллярного аппарата и саркоплазматического ретикулума, а также достаточно высокая проницаемость плазматической мембраны для ионов натрия и в связи с этим невысокий мембранный потенциал покоя. В процессе эмбрионального развития большая часть этих исходно обладающих автоматией кардиомиоцитов претерпевает определенные структурные изменения, утрачивает способность к автоматии и становится рабочими кардиомиоцитами. Сравнительно небольшая часть эмбриональных кардиомиоцитов сохраняет способность к автоматии, черты строения эмбриональной мышечной ткани и образует проводящую систему сердца.
М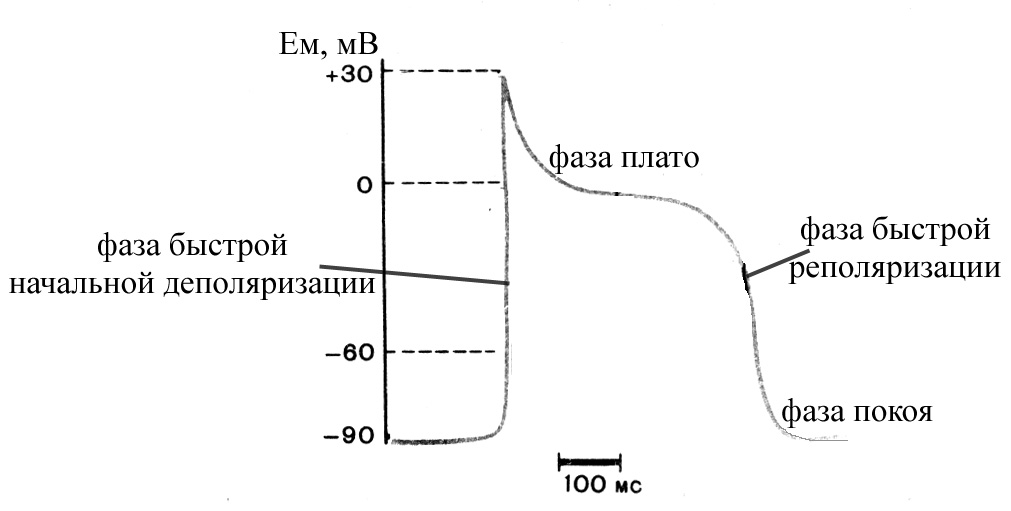 ембранный
потенциал покоя рабочих кардиомиоцитов
создается преимущественно выходящим
калиевым током (обычный ток утечки)
и его значение (-90 мВ) приближается к
калиевому равновесному потенциалу. О
потенциале покоя проводящих кардиомиоцитов
можно говорить лишь условно, поскольку
для них после завершения каждого
потенциала действия характерна спонтанная
диастолическая деполяризация (СДД),
которая и обуславливает их ритмическую
активность. Мембранный потенциал
покоя проводящих кардиомиоцитов гораздо
ниже (-50 ÷ -60 мВ) такового рабочих,
поскольку их мембрана в покое намного
лучше проницаема для натрия. В связи
с этим в проводящих кардиомиоцитах в
создании потенциала покоя, наряду с
выходящим калиевым током,
участвует и довольно выраженный
входящий натриевый ток (подобно
выходящему калиевому току является
током утечки), уменьшающий степень
поляризации мембраны, обусловленной
выходом калия из клетки.
ембранный
потенциал покоя рабочих кардиомиоцитов
создается преимущественно выходящим
калиевым током (обычный ток утечки)
и его значение (-90 мВ) приближается к
калиевому равновесному потенциалу. О
потенциале покоя проводящих кардиомиоцитов
можно говорить лишь условно, поскольку
для них после завершения каждого
потенциала действия характерна спонтанная
диастолическая деполяризация (СДД),
которая и обуславливает их ритмическую
активность. Мембранный потенциал
покоя проводящих кардиомиоцитов гораздо
ниже (-50 ÷ -60 мВ) такового рабочих,
поскольку их мембрана в покое намного
лучше проницаема для натрия. В связи
с этим в проводящих кардиомиоцитах в
создании потенциала покоя, наряду с
выходящим калиевым током,
участвует и довольно выраженный
входящий натриевый ток (подобно
выходящему калиевому току является
током утечки), уменьшающий степень
поляризации мембраны, обусловленной
выходом калия из клетки.
Рис. 33. Схема потенциала действия рабочего кардиомиоцита (по Косицкому Г.И., 1985)
В потенциале действия (ПД) рабочих кардиомиоцитов выделяют следующие фазы:
фаза быстрой начальной деполяризации (продолжительность 1-2 мс), обусловлена быстрым входящим потенциалзависимым натриевым током, возникает вследствие активации (открытия быстрых активационных ворот) быстрых потенциалзависимых натриевых каналов; характеризуется большой крутизной нарастания, поскольку обуславливающий ее ток обладает способностью к самообновлению. Потенциалзависимые натриевые каналы имеют два типа ворот (активационные и инактивационные), срабатывающие на деполяризацию с разной скоростью (активационные быстро открываются, а инактивационные гораздо медленнее закрываются). В связи с этим потенциалзависимые натриевые каналы сравнительно быстро открываются при начинающейся деполяризации мембраны рабочего кардиомиоцита (при мембранном потенциале (МП) -60÷-70 мВ бóльшая их часть уже оказывается открытой), но при этом сравнительно быстро и инактивируются (при МП -40 мВ бóльшая их часть уже инактивируется). Специфическим блокатором электровозбудимых натриевых каналов является тетродотоксин. Местные анестетики оказывают угнетающее действие на электровозбудимые натриевые каналы, что проявляется в замедлении восстановления их проницаемости после инактивации и, как следствие этого, удлинении периода абсолютной рефрактерности без изменения общей продолжительности потенциала действия.
фаза плато ПД, обусловлена потенциалзависимым медленным входящим кальциевым током (этот ток является медленным в связи с медленным открытием потенциалзависимых кальциевых каналов). Начальная деполяризация мембраны, вызванная входящим натриевым током, приводит к открытию медленных кальциевых каналов, через посредство которых ионы кальция по концентрационному градиенту входят внутрь кардиомиоцита; эти каналы в гораздо меньшей степени, но все же проницаемы и для ионов натрия. Медленные потенциалзависимые кальциевые каналы активируются деполяризацией мембраны (активация начинается при МП, равном -40÷ -30 мВ, а полная активация наступает при МП 0÷+10 мВ), имеют два типа ворот (активационные и инаткивационные), инактивируются также деполяризацией. Специфическими блокаторами электровозбудимых кальциевых каналов являются ионы кадмия, а также некоторые органические вещества (верапамил, нифедипин, дилтиазем). Вход кальция и отчасти натрия в кардиомиоцит через медленные кальциевые каналы несколько деполяризует его мембрану (но гораздо слабее, чем предшествующий этой фазе быстрый входящий натриевый ток). В эту фазу быстрые натриевые каналы, обеспечивающие фазу быстрой начальной деполяризации мембраны, инактивируются, и клетка переходит в состояние абсолютной рефрактерности (потенциалзависимые натриевые каналы остаются инактивированными до тех пор, пока мембрана не реполяризуется до уровня -40 мВ). В период фазы плато потенциала действия происходит и постепенная активация потенциалзависимых калиевых каналов II типу (канали із внутрішнім вирівнюванням), які пропускають іони калію всередину клітини, але перешкоджають їхньому виходу в позаклітинну рідину і лише при низьких значеннях мембранного потенціалу починають пропускати їх за межі клітини. Эта фаза является самой длительной фазой ПД (составляет 0,27 с при общей длительности ПД 0,3 с), в результате чего кардиомиоцит большую часть времени в период генерации ПД находится в состоянии абсолютной рефрактерности. Причем длительность одиночного сокращения миокардиальной клетки (около 0,3 с) примерно равна таковой ПД, что вместе с продолжительным периодом абсолютной рефрактерности делает невозможным развитие тетанического сокращения сердечной мышцы, которое было бы равнозначно остановке сердца. Следовательно, сердечная мышца способна к развитию только одиночных сокращений. Кроме того, длительный период абсолютной рефрактерности в сердечных мышечных клетках предохраняет миокард от слишком быстрого повторного возбуждения, которое могло бы нарушить нагнетательную функцию сердца. В частности, фаза рефрактерности препятствует круговому движению возбуждения по миокарду, поскольку рефрактерный период кардиомиоцитов больше, чем время распространения возбуждения по предсердиям или желудочкам. В связи с этим каждая очередная волна возбуждения из синоатриального узла, охватив полностью весь миокард, угасает (обратный вход ее не возможен, поскольку сердце находится в состоянии рефрактерности). Отмеченное обстоятельство ограничивает частоту сердечных сокращений и делает возможным нахождение миокарда в состоянии общей паузы после каждой систолы желудочков до момента возникновения очередного возбуждения в синоатриальном узле
фаза быстрой реполяризации, обусловлена быстрым выходящим потенциалзависимым калиевым током, в период этой фазы медленные кальциевые каналы постепенно инактивируются, что ускоряет процесс реполяризации мембраны (поскольку резко ослабевает входящий кальциевый ток, деполяризующий мембрану). Потенциалзависимые калиевые каналы имеют только один тип ворот (активационные), срабатывающие на деполяризацию открытием, а на реполяризацию – закрытием. Активация этих каналов начинается уже при МП -40÷-50 мВ, а полная активация достигается при МП +10 мВ (в момент фазы плато ПД). Реполяризация мембраны сопровождается постепенным закрытием потенциалзависимых калиевых каналов (полное их закрытие может длиться до 1 с и поэтому, как правило, продолжается еще и в начале каждого следующего ПД) и реактивацией быстрых натриевых каналов (их возвратом к исходному состоянию, характерному для покоя), что приводит к постепенному восстановлению возбудимости миокардиальной клетки. В связи с отмеченным в эту фазу миокардиоцит находится в состоянии относительной рефрактерности.
Реполяризация мембраны рабочего кардиомиоцита приводит к восстановлению мембранного потенциала до уровня, характерного для состояния покоя (-90 мВ), и клетка продолжает пребывать в этом состоянии до момента прихода очередного нервного импульса. Амплитуда потенциала действия рабочего кардиомиоцита составляет около 120 мВ (реверсия 30 мВ).
Потенциал действия проводящих кардиомиоцитов характеризуется рядом отличительных черт от такового в рабочих клетках. Во-первых, он имеет гораздо меньшую амплитуду (50-60 мВ, реверсия составляет 0÷+10 мВ). Во-вторых, фаза его начальной деполяризации характеризуется меньшей крутизной нарастания, а фаза плато ПД – гораздо меньшей длительностью по сравнению с таковыми в рабочей миокардиальной клетке.
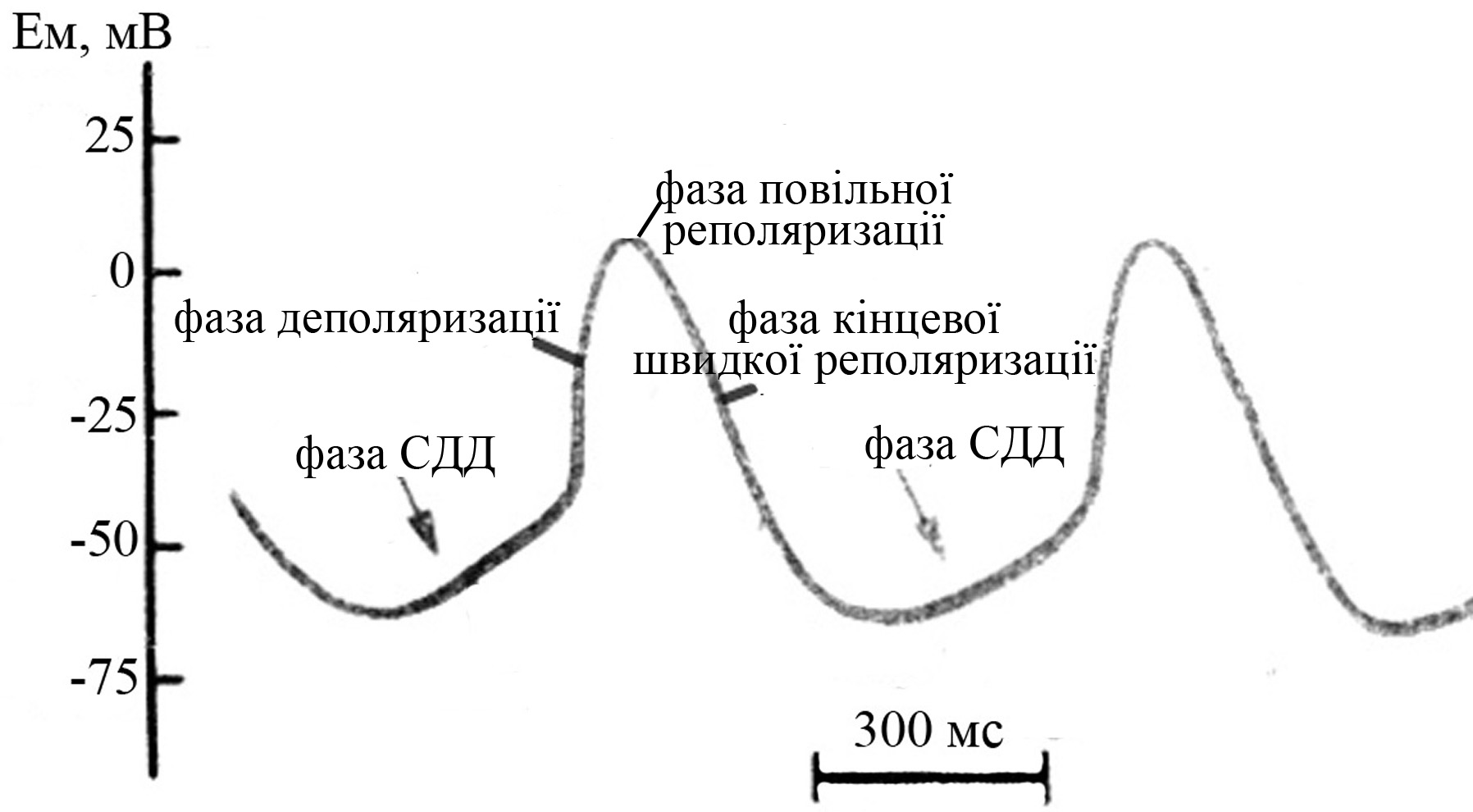
Рис. 34. Схема потенциала действия проводящего кардиомиоцита (по Косицкому Г.И., 1985)
В потенциале действия проводящих кардиомиоцитов выделяют следующие фазы:
фаза спонтанной диастолической деполяризации (СДД), возникает вскоре после возврата МП к исходному уровню после предшествующего возбуждения, обусловлена:
входящим натриевым током утечки (мембрана проводящих кардиомиоцитов в покое хорошо проницаема для ионов натрия)
отчасти быстрым входящим потенциалзависимым натриевым током (потенциалзависимые натриевые каналы в пейсмекерных клетках в большинстве своем инактивируются при достижении мембранным потенциалом уровня -40 мВ, в связи с чем принимают участие в СДД лишь в ее начале). Основная же роль потенциалзависимых натриевых каналов в пейсмекерных клетках состоит в обеспечении восстановления их мембранного потенциала после гиперполяризации, вызванной действием ацетилхолина или других гуморальных факторов, а также противодействии гиперполяризующим процессам со стороны более поляризованных рабочих кардиомиоцитов на проводящие кардиомиоциты
некоторым уменьшением калиевой проницаемости мембраны. В частности, потенциалзависимые калиевые каналы, обеспечивающие реполяризацию мембраны кардиомиоцита, характеризуются очень длительной инактивацией, которая продолжается уже после ПД в момент СДД следующего ПД. Постепенное же полное закрытие этих каналов приводит к резкому ослаблению выходящего потенциалзависимого калиевого тока, что также способствует некоторой деполяризации мембраны кардиомиоцита в момент начинающейся очередной СДД
отчасти медленным входящим потенциалзависимым кальциевым током, включается во время последней 1/3-1/4 СДД і здійснюється переважно за участю кальцієвих каналів Т-типу (від англ. transient – тимчасовий). Потенциалзависимые медленные кальциевые каналы начинают активироваться деполяризацией при МП -40 мВ, они гораздо хуже, но все же проницаемы и для ионов натрия
фаза деполяризации, обусловлена медленным входящим потенциалзависимым кальциевым током через кальцієві канали L-типу (від англ. long-lasting – довготривалий). Кальциевый ток является гораздо более медленным, чем потенциалзависимый натриевый ток), в связи с чем отличается гораздо меньшей крутизной нарастания и большей длительностью, чем соответствующая фаза в рабочих кардиомиоцитах, а также меньшей амплитудой. Причем для полного восстановления от инактивации потенциалзависимых медленных кальциевых каналов требуется несколько сотен миллисекунд, и реактивация кальциевых каналов продолжается даже после завершения ПД (уже в начале фазы СДД следующего ПД)
фаза медленной реполяризации, обусловлена продолжающимся, но гораздо более слабым, чем в предыдущей фазе, медленным потенциалзависимым кальциевым током на фоне начавшейся активации калиевых каналов. Уже во время предшествующей фазы деполяризации проводящих кардиомиоцитов медленные кальциевые каналы начинают частично инактивироваться (при МП около -20 мВ), но поскольку их инактивация происходит очень медленно, то еще в течение значительного времени часть этих каналов остается открытой, обеспечивая фазу медленной реполяризации. Данная фаза в проводящих кардиомиоцитах, особенно синоатриального и атриовентрикулярного узлов, отличается гораздо меньшей продолжительностью по сравнению с фазой плато в рабочем миокардиоците
фаза быстрой реполяризации, подобно таковой в рабочих кардиомиоцитах, обусловлена выходящим потенциалзависимым калиевым током. По ходу этой фазы возбудимость проводящих кардиомиоцитов постепенно восстанавливается, а по достижении мембранным потенциалом величины -50 ÷ -60 мВ, начинается новая СДД. При очень частой генерации нервных импульсов в синоатриальном узле и соответственно в других проводящих и рабочих кардиомиоцитах сердца, медленно закрывающиеся по мере реполяризации мембраны калиевые каналы оказываются частично открытыми не только в начале СДД каждого следующего потенциала действия, но и в момент фазы реполяризации, ускоряя тем самым реполяризацию каждого следующего ПД и укорачивая его продолжительность, что способствует еще большему возможному увеличению частоты сердечных сокращений. Селективним блокатором потенціалзалежних калієвих каналів робочих та провідних кардіоміоцитів є аміодарон, введення якого супроводжується блокадою частини електрозбудливих калієвих каналів міокардіальних клітин і, як наслідок, збільшенням тривалості як фази їх реполяризації, так і саме потенціалу дії, але при цьому не виникає загрози повної зупинки серця, яка може виникнути після введення великих доз блокаторів потенціалзалежних натрієвих або кальцієвих каналів внаслідок повного блокування генерації нервових імпульсів справжніми пейсмекерами. Здатність аміодарону блокувати потенціалзалежні калієві канали, збільшуючи тим самим тривалість потенціалу дії провідних кардіоміоцитів та відповідно зменшуючи частоту їх імпульсації, обумовила використання цього препарату в клінічній практиці для лікування тахікардій, у тому числі екстрасистолій та пароксизмальної тахікардії.
Длительность потенциала действия и соответственно рефрактерного периода проводящих кардиомиоцитов увеличивается по мере перехода от синоатриального узла к волокнам Пуркинье, и самой продолжительной она является в окончаниях волокон Пуркинье, которые благодаря этому выступают в качестве частотного фильтра, не пропуская к рабочему миокарду желудочков слишком частые импульсы от эктопических очагов возбуждения предсердий. Причем длительность ПД волокон Пуркинье превышает таковую не только вышерасположенных структур проводящей системы сердца, но и рабочих кардиомиоцитов желудочков.
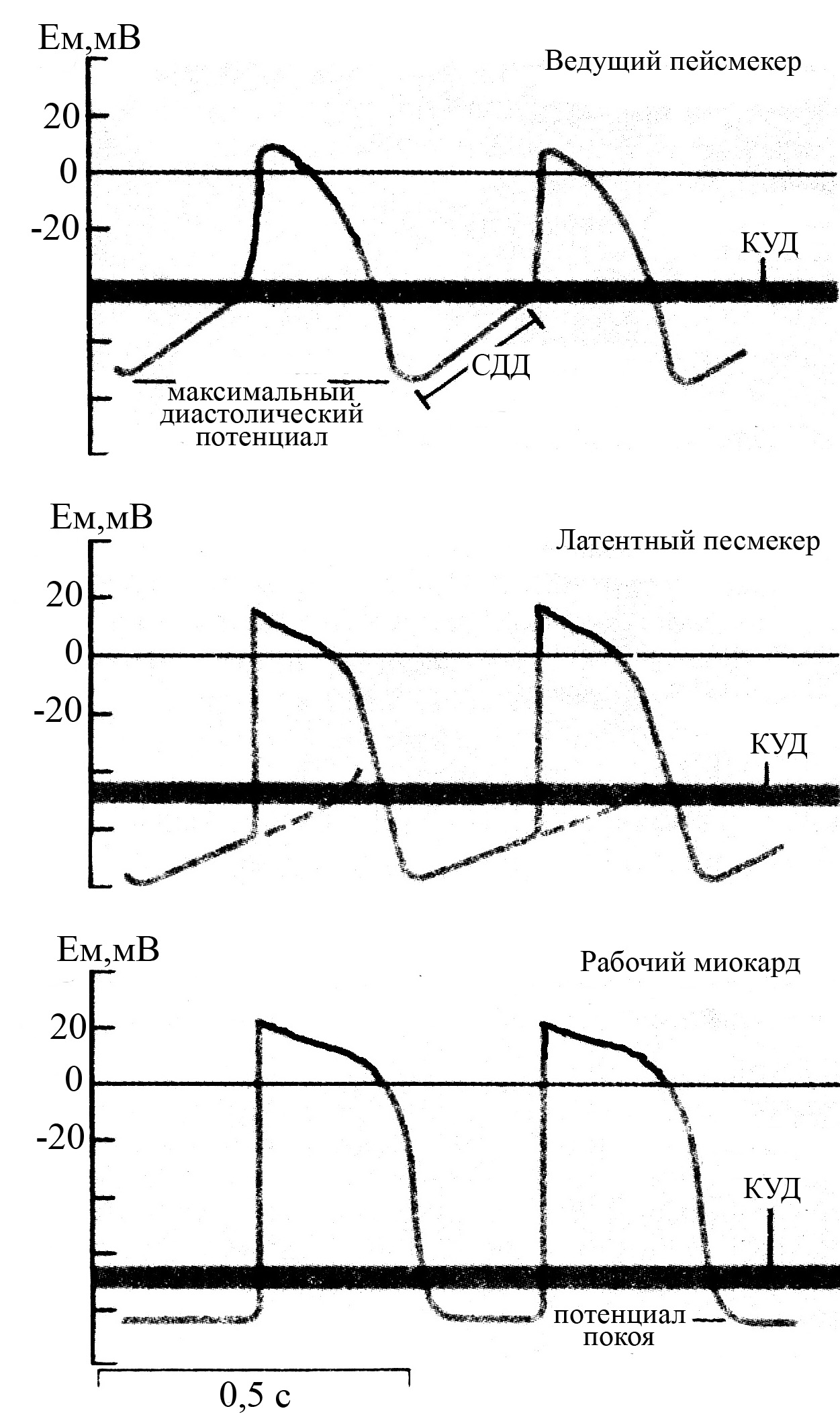
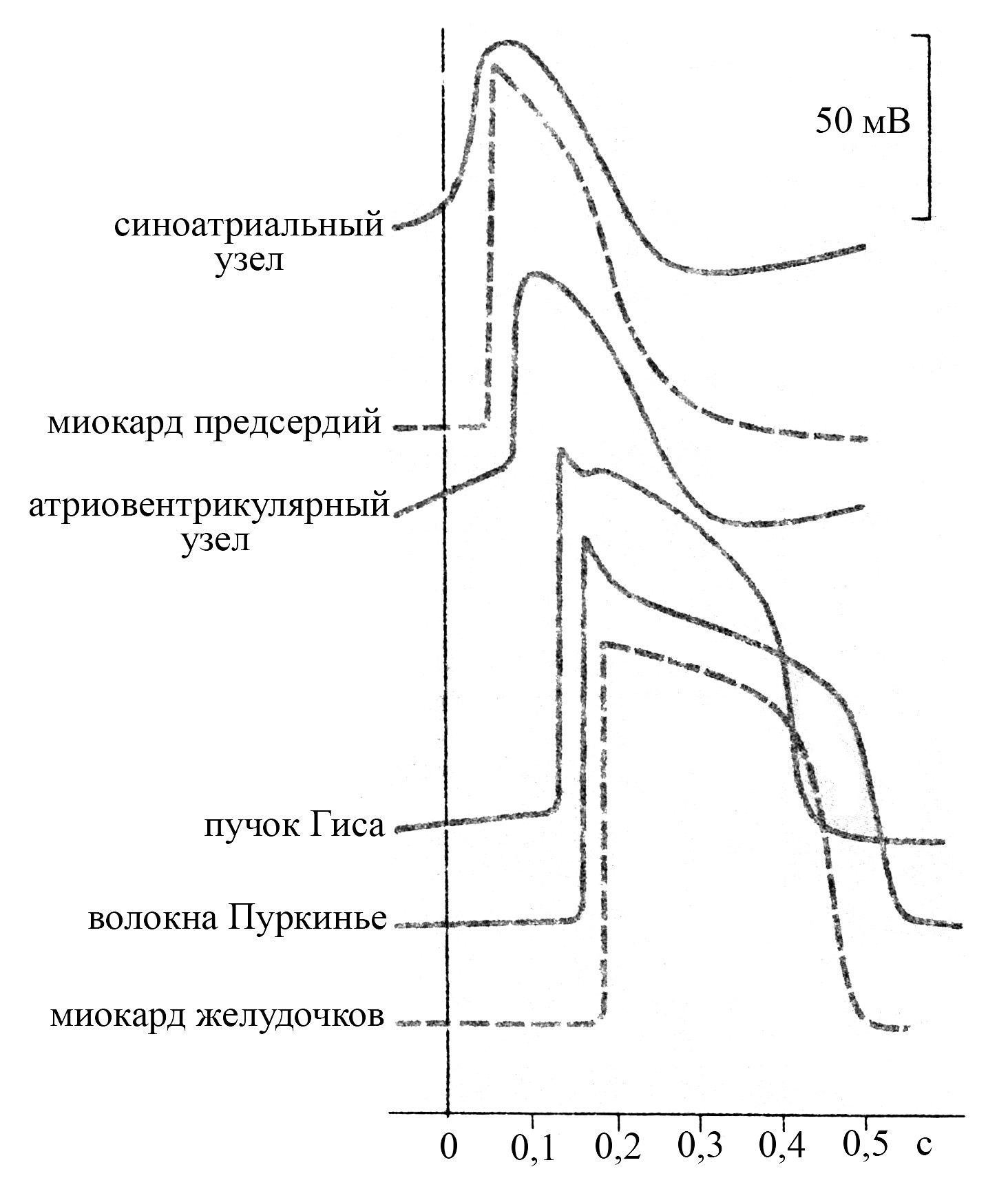
Рис. 36. Типичная конфигурация
потенциалов действия различных
отделов сердца (по Antoni X., 1996).
Проекция начала потенциала действия
на горизонтальную шкалу
соответствует времени прихода
волны возбуждения в этот отдел
Рис. 35. Конфигурация потенциалов действия
различных отделов проводящей системы и
рабочего миокарда (по Antoni X., 1996)
Крутизна переднего фронта и амплитуда потенциала действия в клетках синоатриального и атриовентрикулярного узлов существенно ниже, чем в остальных отделах проводящей системы сердца, и еще гораздо меньше таковых в рабочем миокарде предсердий и желудочков.
Длительность плато и соответственно абсолютного рефрактерного периода в рабочих миокардиоцитах предсердий короче по сравнению с кардиомиоцитами желудочков.
Скорость и крутизна спонтанной диастолической деполяризации в проводящих кардиомиоцитах характеризуются самыми большими величинами в клетках синоатриального узла, а по мере удаления от синоатриального узла эти параметры уменьшаются, что и приводит к урежению частоты нервных импульсов, генерируемых латентными пейсмекерами.