

IX. Органи травлення. Походження
Травна система - це сукупність органів травлення у тварин і людини, що забезпечує організм необхідною енергією та будівельним матеріалом для відновлення клітин і тканин, які безперервно руйнуються в процесі життєдіяльності.
У просто організованих тварин травної системи немає. Для найпростіших характерне внутрішньоклітинне травлення. У найпримітивніших багатоклітинних тварин травлення здійснюється окремими органами: у губок - хоаноцитами й пінакоцитами, у безкишечних війчастих червів - травними клітинами паренхіми. Травна система безхребетних досить різноманітна. У кишковопорожнинних вона представлена гастральною порожниною, висланою ентодермою. Це сліпий мішок, що відкривається назовні єдиним ротовим отвором. У немертин, первиннопорожнинних червів та інших безхребетних, поряд з передньою та середньою кишкою є ще й задня ектодермальна кишка з анальним отвором. Найбільшої складності травна система досягає в членистоногих: середня кишка павукоподібних і ракоподібних має велику травну залозу. У павукоподібних, комах, багатоніжок у кишечник відкриваються органи виділення - мальпігієві судини. Деякі безхребетні в процесі еволюції втратили травну систему: стьожкові черви й скреблянки в результаті тривалого ендопаразитичного способу життя, а погонофори - у зв'язку з життям в захисній трубці.
Надзвичайно різноманітне диференціювання кишечника в хордових тварин. Насамперед, на відміну від членистоногих, майже весь кишковий канал ентодермальний, тим часом як ектодермою вислані лише невеликий початковий і дуже малий кінцевий його відділи. Найхарактернішою особливістю є, однак, більш чи менш значне перетворення передньої частини кишечника у своєрідний орган дихання.
Філогенетичний розвиток у хребетних являє собою яскравий приклад поступової диференціації простого органа - прямої кишечної трубки - в складну систему органів, що виконують окремі функції травлення.
188
Цей процес диференціації спостерігається також в ембріональному періоді розвитку. Первинним зачатком травної системи є ентодермальна трубка, замкнена спереду; ззаду вона переходить у нейрокишечний канал. Ротова порожнина розвивається вп'ячуванням ектодерми (stomodeum), яке проривається в кишечник. Подібне ектодермальне вп'ячування (proctodeum) в задній частині трубки утворює анальний отвір або клоаку.
У результаті диференціювання травний тракт утворює таку систему органів (мал.81):
1) ротова порожнина - орган захоплення їжі, що містить зуби, язик, ротові залози;
2) глотка - виконує дихальну функцію, є місцем виникнення важливих залоз внутрішньої секреції - щитоподібної (gl.thyreoidea), зобної (gl.thymus) й частини гіпофіза (gl. hypophysis);
3) стравохід;
4) шлунок - орган хімічної переробки їжі;
5) дванадцятипала кишка - важливий відділ кишечника, де відбувається перетравлення жирів, білків та вуглеводів;
6) тонкі кишки, де відбувається всмоктування поживних речовин;
7) сліпа кишка, більш характерна для вищих хребетних;
8) товста кишка - резервуар для кінцевого відокремлення неперетрав-лених решток їжі;
9) пряма кишка, де формуються фекальні маси;
10) анальний отвір.
М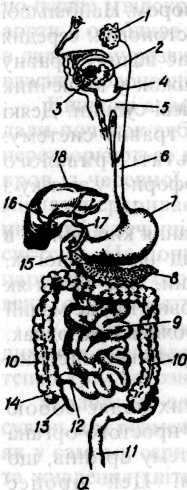 ал.81.
Схема будови органів травлення: 1
-
привушна,
2
- підщелепна,
З -
під'язикова
слинні залози; 4
- надгортанник;
5
- вхід
у гортань; 6
- стравохід;
7
— шлунок;
8
- підшлункова
залоза; 9
- топка
кишка; 10
- товста
кишка; 11
- пряма
кишка 12
- клубова
кишка;
ал.81.
Схема будови органів травлення: 1
-
привушна,
2
- підщелепна,
З -
під'язикова
слинні залози; 4
- надгортанник;
5
- вхід
у гортань; 6
- стравохід;
7
— шлунок;
8
- підшлункова
залоза; 9
- топка
кишка; 10
- товста
кишка; 11
- пряма
кишка 12
- клубова
кишка;
13 - червоноподібний відросток (апендикс),
14 - сліпа кишка, 15 - дванадцятипала кишка; 16 - жовчний міхур і його протока (17);
18 - печінка.
189
Ротова порожнина
Ротовий отвір у хребетних, як правило, розміщається на передньому кінці тіла, але буває більш чи менш зсунутий на черевний бік (селахії, осетрові). Ротова порожнина вислана багатошаровим плоским епітелієм, який інколи ороговіває. У риб і наземних хребетних спереду й з боків ротова порожнина обмежена щелепами (і зубами), знизу -м'язовим дном. Покрівля її зміцнена кістками дна черепа, що утворюють первинне піднебіння й хоани, які відкриваються в ротову порожнину. В амніот ротова порожнина поділяється на верхній (дихальний) і нижній (вторинна ротова порожнина) відділи завдяки розвитку твердого кісткового піднебіння. Від його заднього краю в ссавців відходить м'яке піднебіння, що відокремлює ротову порожнину від глотки. На піднебінні птахів, а ще частіше ссавців, виникають сосочки, поперечні валики або піднебінні пластинки, покриті роговим шаром. Це сприяє перетиранню їжі язиком. У беззубих китів роговий шар цих валиків дуже розростається, утворюючи китовий вус.
У зв'язку з розвитком м'яких рухливих губів у ссавців між губами й зубами є порожнина - пристінок рота, яка в багатьох мавп та гризунів утворює защічні мішки.
У ротовій порожнині хребетних розвивається ряд органів спеціального призначення: зуби, язик, ротові залози.
Зуби - це кісткові утвори, розміщені в ротовій порожнині більшості щелепноротих хребетних, які служать для захоплення, утримання, а інколи й роздріблення їжі. У процесі філогенезу тварин зуби виникли з плакоїдних лусок шкірного походження. Закладаються зуби в період зародкового розвитку у вигляді епітеліальної складки - зубної пластинки із зачатками окремих зубів (мал.82).
М ал.
82. Розвиток
зуба ссавця (з його замісником) (за
Гертвігом): 1
- зубна
пластинка, 2
- емальовий
орган, 3
- м
'якуш зуба, 4
-
зачаток
постійного зуба.
ал.
82. Розвиток
зуба ссавця (з його замісником) (за
Гертвігом): 1
- зубна
пластинка, 2
- емальовий
орган, 3
- м
'якуш зуба, 4
-
зачаток
постійного зуба.
190
Мезенхімні клітини дерми утворюють під кожним зачатком згущення (зубні сосочки), над якими епітеліальні клітини зубної пластинки нависають у вигляді ковпачка, так званого емальового органа. Внутрішній шар клітин зубної пластинки бере участь в утворенні шару емалі, що складається з призматичних волокон, які стоять вертикально до поверхні зуба. Ці волокна покриті тонким шаром органічної речовини - зубною кутикулою. Зовнішні клітини зубного сосочка -одонтобласти - утворюють дентин, що має призматичну будову з численними гіллястими канальцями, в які проникають відростки одонтобластів. Нарешті, у ссавців основа зуба вкривається ззовні шаром кісткової речовини, яка має назву цементу. Всередині зуба є порожнина, заповнена м'якушем (пульпою). Канал відкривається в кінці кореня зуба отвором, через який в порожнину зуба проникають кровоносні судини й нервові закінчення.
У більшості хребетних зуби в міру зношування замінюються новими. Зміна їх відбувається впродовж усього життя. Таку необмежену зміну зубів називають полі-фіодонтизмом (більшість хребетних, крім ссавців). У більшості ссавців спостерігаються лише дві послідовні зубні системи (молочна й постійна) - діфіодонтизм. У деяких випадках у ссавців прорізається лише одне покоління зубів (неповнозубі, китоподібні) - монофіодонтизм.
У більшості риб, плазунів і в земноводних усі зуби однакової форми, кількість їх буває дуже великою. Це гомодонтна система. Деякі риби, плазуни і, як правило, ссавці мають зуби різної форми, які виконують різні функції (гетеродонтна система). У деяких випадках зубна система редукується і тоді розвивається роговий дзьоб або інші утвори, які функціонально її замінюють (черепахи, птахи, клоакові ссавці, беззубі кити).
У круглоротих розвиваються особливі рогові зуби, не гомологічні зубам щелепноротих. Розміщуються вони в ротовій лійці та на язику, являють собою горбки зроговілого епітелію.
У риб зуби розміщуються в м'яких тканинах і на кістках ротової порожнини, на зябрових дугах (глоткові зуби ). Зазвичай вони численні та дрібні. У селяхій вони порівняно великі й бувають ланцетоподібними, зазубреними або у вигляді плоских пластинок.
191
У дводишних риб зуби закладаються у вигляді окремих пластинок, а потім зростаються між собою, утворюючи великі зубні пластинки.
В амфібій є багато дрібних конічних зубів, що сидять на щелепних, передщелепних, зубних, а іноді й на пластинчастих кістках, на лемешах та піднебінних кістках. У деяких видів вони редукуються.
Зуби плазунів звичайно простої конічної форми, але іноді спостерігається диференціювання. Особливо воно характерне для викопних теріодонтів. Зуби приростають або збоку до внутрішньої поверхні щелепних кісток (плевродонтні), або своєю основою до краю щелеп (акродонтні), або сидять в альвеолах (текодонтні). В отруйних змій у верхній щелепі розвиваються особливі зуби, що мають канал, зв'язаний з отрутною залозою - видозмінена слинна залоза. Зміна зубів у плазунів необмежена.
У сучасних птахів зубна система редукована й заміщена функціонально роговим дзьобом. Викопні птахи (археоптерикс, гесперорніс, іхтіорніс та ін.) мали численні конічні зуби, що сиділи в деяких форм в окремих комірках.
У ссавців зубна система текодонтна і, як правило, високо диференційована. Передні зуби - різці (incisivi) долотоподібної форми, служать для захоплення та розрізання їжі. У слонів з них утворюються бивні. Передніх зубів буває від однієї до п'яти пар. За різцями йдуть ікла - (саnіnі) конусоподібні, служать для захоплення й розривання здобичі (в хижаків), є засобом захисту (у всеїдних копитних, деяких ластоногих і китоподібних). Задні зуби - кутні (до 8 пар) -мають складну форму й служать для перетирання їжі. Серед них розрізняють псевдокутні (praemolares) й справжні кутні (molares), які мають два або більше коренів.
Форма та кількість зубів у різних видів ссавців більш чи менш постійні й служать важливою систематичною ознакою.
Процес поступового ускладнення форми кутніх зубів тепер вичерпно простежений на викопних формах. Колишня думка, що кутні зуби ссавців утворились через злиття певної кількості простих конічних зубів, поступилася місцем так званій тритуберкулярній теорії палеонтологів Копа й Осборна. За цією теорією складна форма кутніх зубів ссавців розвинулась шляхом поступового перетворення простих конічних зубів, як це іноді спостерігається і в рептилій, особливо у викопних теріодонтів.
192
Спочатку був ряд конічних зубів (гаплодонтна стадія), які у верхній та нижній щелепі чергуються, як це спостерігається й тепер у хижих та багатьох інших ссавців (мал.83).
М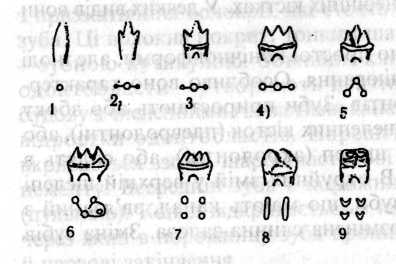 ал.
83. Схема
утворення основних форм кутніх зубів
ссавців: І -
гаплодонтна,
2,З
-
протодонтна,
4-тризубчаста,
5
- тригорбкувата,
ал.
83. Схема
утворення основних форм кутніх зубів
ссавців: І -
гаплодонтна,
2,З
-
протодонтна,
4-тризубчаста,
5
- тригорбкувата,
6 -гострогорбкувата,
7 - чотиригорбкувата,
8 - складчаста,
9 - ямкувата.
Навколо основи конічного зуба був кільцевий валик, від якого утворювались додаткові вершини зуба. Спочатку попереду й позаду від головної вершини розвинулись невеликі додаткові вершини (протодонтна форма). Ці додаткові вершини наростали до розміру головної, утворився зуб з трьома однаковими зубцями в одній площині (тризубчаста форма). Додаткові вершини однієї щелепи входили в проміжок між головною й додатковою вершинами другої щелепи.
Потім, за Осборном, головна вершина змістилась в один бік, а додаткові - в другий. Тепер три вершини зуба розміщаються по кутах трикутника (тритуберкулярна форма). За іншими даними (Грегорі) власне переміщення вершин не було, а тригорбкуватий тип зуба розвинувся самостійно з того ж простого конічного зуба. Така форма зубів спостерігається і в деяких сучасних сумчастих, комахоїдних, напівмавп, рукокрилих.
При подальшому ускладненні спочатку на зубах нижньої щелепи, а потім і на зубах верхньої щелепи розвивається ззаду біля основи коронки виступ (я'ятка) з горбком на ньому. Такі зуби зустрічаються в багатьох сучасних ссавців, і ця форма лежить в усякому разі в основі різних типів комахоїдних і хижих. Якщо вершини такого зуба сполучені між собою гострим ріжучим краєм, то зуб дістає назву ріжучого (секодонтний).
193
Є ще гострогорбкуваті й тупогорбкуваті (бунодонтні) форми зубів.
Подальше ускладнення будови зуба виявляється в тому, що між головними горбами розвиваються додаткові. Нижній горб на п'ятці може досягти розміру інших трьох горбів, що спускаються до рівня горба п'ятки, і тоді ми маємо чотири-горбкуватий (квадритуберкулярний) тип зубів. З такого зуба виводяться і складчасті (лоходонтні) зуби, характерні для коней, тапірів. З чотиригорбкуватого зуба виводяться також ямкуваті (селенодонтні) зуби жуйних через зміну форми горбів, які витягуються й згинаються під кутом (мал.83).
Нарешті необхідно ще зазначити, що при найвищому диференціюванні зуби травоїдних ссавців набувають дедалі тривалішого росту, корені в них не розвиваються і, відповідно, змінюється форма зубів - короткі, брахіодонтні зуби з обмеженим ростом перетворюються в високі, циліндричні, гіпселодонтні зуби з тривалим ростом, причому горбки витягуються в довгі призми, а в проміжках між призмами відкладається цемент. Палеонтологія дає нам чудові ряди форм, що ілюструють цей процес.
Отже зуби хребетних, що виникли як суто шкірні захисні утвори (плакощна луска), набули значних розмірів на щелепах, де вони відіграють роль допоміжних органів схоплювання й утримування здобичі. Уже в примітивних хребетних їх форма змінюється у зв'язку із зміною характеру їжі. У ссавців зуби значно диференціюються, так що розвиваються принаймні три основні типи зубів, які розрізняються і формою, і функцією. Зуби сидять в окремих комірках, основа їх звужується, утворюючи корінь. Окремі зуби збільшуються в розмірах, кутні набирають складної форми, але кількість зубів зменшується, одночасно зменшується кількість змін (не більше двох поколінь зубів). Із збільшенням тривалості функціонування таких зубів, особливо при значному їх стиранні, тривалість росту збільшується, так що іноді зуби ростуть впродовж усього життя. Ця еволюція зубної системи мабуть зв'язана з набуттям здатності попереднього розжовування їжі, що у свою чергу привело до більш швидкого й досконалого її перетравлення й засвоювання, яке характеризує енергійніший обмін речовин та інтенсивнішу життєдіяльність, наприклад, таку, як у ссавців.
194
На дні ротової порожнини в хребетних є непарний виступ - язик (lingua). У риб язик утворений складкою слизової оболонки, не має власної мускулатури (за винятком дводишних) і рухається разом з усім вісцеральним скелетом завдяки рухам під'язиково-зябрового апарату. У наземних хребетних він набуває значення рухомого органу, що служить не тільки для втримування й переміщення їжі в ротовій порожнині, а й для її захоплювання, випробування, подрібнення, ковтання, а також для відокремлення й замикання дихальних шляхів. У всіх наземних хребетних розвивається мускулатура язика, що являє собою похідне під'язикової парієтальної мускулатури. М'яз, що висуває язик уперед (підборідно-язиковий), і той, що втягує язик назад (під'язиково-язиковий) відокремились від підборідно-під'язикового та грудинно-під'язикового м'язів. Від цих же м'язів, мабуть, відокремилась і мускульна маса самого язика. Відповідно до свого походження мускулатура язика іннервується язикового-глотковим нервом.
Будова язика земнововодиих надзвичайно різноманітна. У деяких випадках він являє собою орган, побудований трохи складніше, ніж у риб. Проте, це спрощення, мабуть, вторинне. У хвостатих та безногих земноводних мускулистий язик прикріпляється до дна ротової порожнини і здатний більш чи менш значно висуватись назовні, а в більшості безхвостих він прикріпляється переднім кінцем до нижньої щелепи, а його вільний задній кінець лежить на дні ротової порожнини і при ловінні здобичі викидається вперед з рота. Залозисті клітини слизової оболонки виділяють липкий секрет, що забезпечує прилипання здобичі до язика.
Язик вищих хребетних являє собою морфологічно складніший утвір, якщо спиратися на дані історії розвитку. Він розвивається з непарного зачатка, що лежить у ділянці другої та третьої зябрових дуг, до якого приєднується потім ще пара валків, що лежать по боках спереду під'язикової дуги. Між цими зачатками розміщається закладка щитоподібної залози. Задній зачаток дає початок кореневі язика, передні парні зачатки згодом зливаються між собою й дають початок тілу язика.
В іннервації язика в рептилій бере участь, крім двох раніше названих нервів, ще трійчастий нерв. Форма й ступінь
195
рухомості язика плазунів надзвичайно різні. У черепах і крокодилів він малорухомий, а в більшості ящірок та змій має видовжену форму, на кінці глибоко роздвоєний і відзначається дуже великою рухливістю й чутливістю. Разом з якобсоновим органом він служить і як хеморецептор. У цьому випадку в основі язика є кільцева складка, що утворює піхву язика. Під'язиковий апарат утворює скелет язика, служить місцем прикріплення м'язів, що рухають язик. Особливо довгий розширений на кінці язик хамелеонів може блискавично викидатись і служить для захоплення дрібної рухливої здобичі.
Язик птахів малорухливий, покритий роговим епітелієм, досить різноманітний за формою, що пов'язано з характером корму. У зерноїдних птахів язик утримує насіння на краю дзьоба при розкушуванні. У колібрі він звернутий у трубочку, через яку засмоктується нектар. У дятлів язик дуже довгий, на кінці має гачкоподібні шипи, що полегшує добування комах з-під кори.
Язик ссавців розвивається загалом аналогічно з язиком плазунів. Власна мускулатура досягає найбільшого розвитку, так що язик стає надзвичайно рухомим і виконує різноманітні функціі. Мускулатура язика поперечносмугаста; переплетення пучків м'язів забезпечує високу диференційованість рухів. Іннервується язик ссавців трьома нервами: під'язиковим (руховим), язиково-глотковим і язиковим (відчуваючими). Він надзвичайно багатий на залози і має сосочки різної форми й значення, почасти відчуваючі, почасти зроговілі. Зроговіння язика іноді буває досить значним, як наприклад, у хижаків. Дуже довгий і клейкий язик у мурахоїдів.
Складні ротові залози являють собою прогресивні набуття наземних хребетних. Спочатку вони служили для змочування слизової оболонки ротової порожнини, запобігаючи її висиханню, а також для змочування їжі, що полегшувало її ковтання. Згодом у ссавців слина має не тільки слиз, а й ферменти, зокрема амілазу, що діє на крохмаль (полісахарид), розщеплює його до мальтози (дисахарид) і мальтази, що діє на дисахарид і розщеплює його до глюкози (моносахарид). Ферментів, що розщеплюють білки й жири, у слині немає. Реакція слини слабо лужна, що створює сприятливе середовище для дії цих ферментів; у кислому середовиші дія ферментів слини припиняється.
196
У наземних амфібій є звичайно непарна міжносова залоза (gl. internasalis) трубчастої будови й піднебінні залози, що виділяють слизистий секрет. У рептилій, крім цих залоз, є ще особливі під'язикові (gl. sublinguales), губні (gl.labrales) і зубні залози. В отруйних змій саме пара задніх верхньозубних залоз перетворена в складну трубчасту отрутовидільну залозу, але вони прикриті почасти жувальним м'язом, при скороченні якого вміст залози видавлюється та виливається протокою до борозни або каналу отруйного зуба. Набуття отруйного апарату не стало новою еволюційною подією, так як у предків земноводних - вимерлих кистеперих риб були отруйні залози в ротовій порожнині, які виконували приблизно ту ж роль (збільшували ефективність здобування їжі).
Птахи мають слизові залози на піднебінні та під'язикові залози, які особливо розвинені в зерноїдних видів.
У ссавців ротові залози досягають найвищого розвитку. Крім численних дрібних слизових залоз - губних, щічних, піднебінних, язикових, які мабуть, порівнювані з однойменними залозами земноводних і плазунів, є ще кілька великих так званих слинних залоз: під'язикова (gl. sublingualis), задня язикова (gl. submaxillaris), привушна (gl. parotis). 3 цих залоз перші дві, схоже, є результатом диференціації під'язикової залози плазунів, а привушна - новим набуттям ссавців, яке розвинулося із щічних залоз.
ДИФЕРЕННЦІАЦІЯ І СПЕЦІАЛІЗАЦІЯ ПЕРВИННОЇ КИШКИ
Глотка (pharinx) - це ділянка травного тракту, розміщена між ротовою порожниною та стравоходом. Стінки її вислані багатошаровим епітелієм і мають добре розвинену поперечносмугасту мускулатуру. У ділянці глотки розвивається весь зябровий апарат. У нижчих хордових кількість зябрових щілин коливається від двох (сальпи, апендикулярії) до кількох сотень (асцидії). У стінці глотки безщелепних розвивається 5-17 пар зябрових щілин, а в риб - 5-8 пар. У зародків наземних хребетних у глотці закладаються рудименти зябрових щілин – карманоподібні
197
закладки зябрових мішків. З їх останньої пари розвиваються легені. Похідними епітелію глотки є волова й щитовидна залози, а в наземних хребетних ще й навколощитовидні. У риб глотка веде в стравохід, а в наземних хребетних з утворенням легеневого дихання та середнього вуха в глотку окремо відкриваються стравохід, гортань, євстахієві труби. Крім того, в ссавців та деяких плазунів (крокодили) у зв'язку з розвитком твердого кісткового піднебіння у верхній відділ глотки відкриваються хоани. Іннервується глотка язико-глотковим та блукаючим нервами.
Стравохід (oesophagus) - це відділ травного тракту тварин і людини, який іде безпосередньо за глоткою і служить для проведення їжі. Від відділу, що йде за ним, стравохід різко не відмежований (тільки в ссавців встановлюється зовнішня межа), але загалом відрізняється від нього внутрішньою вистилкою з багатошарового епітелію, подібно до глотки та ротової порожнини, а почасти й положенням поза черевною порожниною і, отже, відсутністю серозної оболонки, що особливо характерно для вищих хребетних. У стінках стравоходу немає травних залоз і внутрішній шар епітелію може навіть роговіти. Стінка стравоходу складається зі сполучнотканинного пухкого шару; розташованого під ним м'язового шару з зовнішніх поздовжніх і внутрішніх циркулярних волокон; підслизового шару й слизової оболонки, в яких розміщені залози. Іннервується стравохід симпатичними, спинномозковими й блукаючими нервами.
Шлунок (gaster) - звичайно розширений відділ кишечника, який зовнішньо чітко відмежований від стравоходу тільки в ссавців, але й у цьому випадку межа є до певної міри умовною, бо фізіологічна межа між цими відділами може не збігатися із зовнішньою видимою.
Фізіологічно шлунок характеризується як орган, у якому проходить перша фаза травлення в кислому середовищі. Стінка його вислана слизовою оболонкою з сіткоподібним розміщенням складок. Вона має одношаровий циліндричний епітелій і трубчасті травні залози. У складі шлункового соку переважає пепсин, але входять і ферменти хімозин і ліпаза. Основні функції шлунка - депонування, механічна й хімічна обробка, евакуація їжі в кишечник. Захисна функція шлунка проявляється в бактерицидній і бактеріостатичній дії,
198
пов'язаній з обробкою мікроорганізмів, які поступають з їжею, соляною кислотою та речовиною типу лізоциму. Первинну роль відіграє екскреторна діяльність шлунка -виділення в його порожнину продуктів проміжного обміну. Шлунок зв'язаний з виробленням кров'яних тілець, так як його залози виділяють необхідний для цього процесу "внутрішній фактор" (фактор Касла). Отже, діяльність шлунка тісно пов'язана з підтримкою гомеостазу в організмі, водно-сольовим обміном, функцією нирок, залоз внутрішньої секреції, кровообігом.
Розміри, форма, деталі будови шлунка в усіх класах хребетних змінюються в широких межах, відображаючи кормову спеціалізацію окремих груп.
У круглоротих і деяких риб шлунок не диференційований. У риб він, як правило, підковоподібно зігнутий. Передня його частина, що бере початок від стравоходу, називається кардіальною, а висхідне коліно, що відкривається в 12-типалу кишку - пілоричною. У кардіальному відділі під дією пепсину в кислому середовищі розпочинається перетравлення їжі; в пілоричному відділі їжа обробляється трипсином, що поступає сюди з передньої частини кишечника, зв'язаної з підшлунковою залозою. У деяких риб (коропові, бички) шлунок відсутній: за коротким стравоходом зразу починається кишечник. У таких риб пепсин не виробляється і його функцію виконує трипсин у тонкому кишечнику. В області шлунка в кісткових риб зазвичай розвиваються пілоричні придатки, які служать для збільшення травної поверхні кишечника та нейтралізації їжі при переході її з кислого середовища шлунка в лужне кишечника.
В амфібій та рептилій шлунок відокремлюється більш чітко, особливо в крокодилів, у яких у межах самого шлунка спостерігається подальше диференціювання: за великим мішкоподібним залозистим відділом із значною мускулатурою йде ясно відокремлена пілорична частина.
У птахів подібне диференціювання приводить до повного відокремлення двох відділів переднього трохи здутого залозистого шлунка й відокремленого від нього чіткою перетяжкою мускульного. У багатьох птахів кутикула мускульного шлунка утворює вирости, які компенсують відсутність зубів (разом з проковтнутими камінцями гастролітами, сприяють механічній переробці їжі).
199
У хижих птахів мускульний шлунок тонкостінний; у зерноїдних, комахоїдних, всеїдних - товстостінний; у рибоїдних він дуже малий, а залозистий утворює великий мішок.
Найбільшого розвитку й диференціювання досягає шлунок ссавців. Його розміри, форма, будова змінюються в різних рядах (мал. 84).
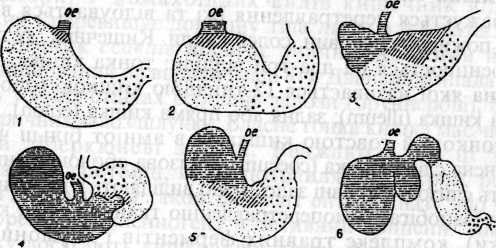
Мал.84. Схема шлунка людини (1), собаки (2), пацюка (3), хом'яка
(4), коня (5), жуйної (6) з позначенням залозистих ділянок
(за Оппелем). Багатошаровий епітелій стравохідного характеру
позначений горизонтальними штрихами; ділянка кардіальних залоз -
косими, донних - пунктиром, пілоричних - хрестиками.
У однопрохідних шлунок має форму простого мішка й позбавлений травних залоз. Однокамерний шлунок хижаків, приматів, комахоїдних формою нагадує реторту; епітелій його стінок має численні травні залози. У жуйних травоїдних ссавців складний шлунок складається з рубця, сітки, книжки й сичуга. Перші три відрізки утворюють так званий передшлунок і вислані багатошаровим епітелієм. Тут немає травних залоз і відбувається лише бактеріальне бродіння з участю симбіонтів, які можуть існувати тільки в нейтральному або слаболужному середовищі. Обробка їжі шлунковим соком відбувається тільки в сичузі, що являє собою справжній шлунок залозистого характеру.
Травні залози шлунка ссавців неоднакові. У відділі, розміщеному ближче до стравоходу, розвинені кардіальні залози; у середньому його відділі - фундальні, а в задньому - пілоричні.
200
Кардіальні залози виділяють слизистий лужний секрет, донні (фундальні) - кислий. У донних залозах розрізняють залозисті клітини двох типів: кубічні - головні клітини, що виділяють проферменти - пепсиноген та прохімозин, і великі округлі покривні клітини, що розсіяні серед головних і виділяють соляну кислоту. Крім того фундальні залози виділяють ліпазу. Пілоричні залози виділяють також пепсиноген і прохімозин.
Кишка власне - це орган травної системи, в якому продовжується перетравлення їжі та відбувається всмоктування поживних речовин, солей і води. Кишечник, як правило, диференціюється на два-три відділи: тонка кишка, передня частина якої називається 12-типалою кишкою (duodenum); товста кишка (illeum); задня або пряма кишка (colon). На межі між тонкою й товстою кишками в амніот більш чи менш розвинена сліпа кишка (соесшп). Слизова оболонка кишечника містить слизові й травні залози, що виділяють слизистий секрет муцин (запобігає самоперетравленню тканин стінок кишечної трубки), комплекс травних ферментів і гормони. Добре розвинена м'язова оболонка, яка здійснює перистальтику кишечника і рух харчових мас. У кишечник поступають ферменти підшлункової залози й жовч. Внутрішня поверхня кишечника збільшується за рахунок розвитку спіральної складки в круглоротих та нижчих риб, або за рахунок подовження самої кишки, яка при цьому утворює більш чи менш значні закрути (петлі). Крім того, збільшення всисної поверхні досягається розвитком складок та ворсинок.
Кишечник круглоротих має вигляд недиференційованої, майже прямої трубки із слабо розвиненим спіральним клапаном (у міног). У нижчих риб (селахій, ганоїдів, дводишних) він має нерідко ще вигляд майже прямої трубки, однак у цьому випадку добре розвинена спіральна складка, яка у вищих риб редукується. З редукцією спіральної складки сама кишка видовжується й утворює численні петлі. Внутрішня стінка кишки має систему складок, на яких іноді є ворсинки. У кишечнику земноводних спостерігається чіткий поділ на тонку й товсту кишки, а в плазунів між ними є складка слизової оболонки (клапан) і невеликий сліпий виступ. У птахів кишечник відносно довгий. Тонка кишка утворює кілька петель і переходить у коротку пряму кишку, що впадає в клоаку.На межі між тонкою й прямою кишками
201
розміщені парні вирости - сліпі кишки, які в більшості видів слабо розвинені. У пташенят розвинений товстостінний сліпий виріст на спинному боці клоаки - фабрицієва сумка, в якій формуються білі кров'яні тільця; у дорослих форм вона редукується. Кишечник довгий у рослиноїдних видів і перевищує довжину тіла в 10 і більше разів; у деяких з них добре розвинені й сліпі кишки, в яких їжа перетравлюється під дією власних ферментів і при участі специфічної мікрофлори. У комахоїдних видів кишечник відносно короткий і перевищує довжину тіла лише в 4-6 разів.
Кишечник ссавців досягає значної довжини й чітко диференційований на відділи (мал. 81). Дванадцятипала кишка містить у своїх стінках особливі залози (бруннерові), близькі до пілоричних залоз шлунка. Власне тонка кишка має численні петлі й ліберкюневі залози. Вважають, що ці залози виробляють серотонін, який, потрапляючи в кров, регулює тонус м'язів судин, а також фермент ентерокіназу, що сприяє перетворенню трипсиногену в активний трипсин. Значного розвитку досягають кишкові ворсинки. Товста кишка часто утворює послідовні виступи, відділені один від одного перехватами, а поздовжня мускулатура обмежена небагатьма мускульними стрічками (taenia), що розділяють ряди виступів. У нижчих однопрохідних ссавців кінцевий відділ кишечника утворює клоаку, в сумчастих вона рудиментарна. У всіх вищих ссавців клоаки немає; пряма кишка з анусом відокремлена від сечостатевого синуса. Довжина кишечника та співвідношення його відділів відповідають складу їжі. Живлення багатими клітковиною кормами супроводжується не тільки загальним видовженням кишечника, а й збільшенням відносних розмірів товстих і, особливо, сліпої кишки, де розвиваються процеси бродіння за участю бактерій, грибків, найпростіших.
Травні залози
Печінка (hepar) - велика залоза тваринного організму, що бере участь у процесах травлення, обміну речовин, здійснює специфічні захисні та знешкоджуючі, ферментативні й видільні функції, спрямовані на підтримку гомеостазу. Тут утворюється жовч, яка емульгує жир.
202
У безхребетних печінка - травна залоза, що являє собою виріст середньої кишки, бере участь в перетравленні та всмоктуванні їжі; у ній відкладаються запасні поживні речовини - жири й вуглеводи.
У хордових печінка - орган, у якому виробляється секрет (жовч), що бере участь у травленні, в ній здійснюються найважливіші процеси, пов'язані з обміном речовин в організмі. У покривників печіяка утворюється як виріст шлунка, тісно прилягає до його стінки й відкривається в нього одним протоком, має вигляд розгалуженої гілочки. У безчерепних вона представлена мішкоподібним печінковим виростом.
У хребетних печінка закладається як черевний виріст середньої кишки, який, деревовидно розгалужуючись, набуває будови трубчастої залози. Просвіти кінцевих трубочок печінки утворюють жовчні капіляри, по яких жовч стікає в більші печінкові протоки. Окремі печінкові протоки об'єднується в загальну жовчну протоку, що відкривається у дванадцятипалу кишку. Як правило, з частини жовчного протоку утворюється жовчний міхур.
Трубчаста будова печінки зберігається впродовж усього життя тільки в деяких круглоротих (міксин). У міног, риб та земноводних трубчаста будова печінки частково порушується, так як між трубочками її виникають поперечні перекладини - анастамози й між ними вростає сполучна тканина з кровонос-ними судинами та нервами. У плазунів, птахів і ссавців добре розвинені анастамози перетворюють трубчасту залозу в сітчасту. У міног і деяких риб печінка -нерозчленований орган, однак у більшості хребетних у ній розвинені ліва й права лопаті, а жовчний міхур завжди зв'язаний з останньою. У деяких амніот (особливо в ссавців) обидві лопаті можуть бути часточковими. Форма печінки деякою мірою залежить від форми тіла тварини (мал. 85).
Маса печінки більша в плотоїдних видів порівняно з травоїдними. Особливої уваги заслуговує печінка хрящових риб. Ця трилопатева залоза складає від 14 до 25 % маси тіла. У ній накопичуються великі запаси жиру (в гігантської акули до 70 % від маси печінки; у деяких скатів - до 60 %), що визначає її роль не тільки як депо скритих резервів, а й як гідростатичного органа, що забезпечує плавучість тіла.
203
Фізіологічне значення печінки визначається тим, що всі речовини, які всмоктуються з кишечника в кров, зазнають хімічних змін у печінці. Вона бере участь у підтриманні динамічної рівноваги багатьох речовин плазми крові, інактивує деякі гормони, тією чи іншою мірою бере участь в обміні білків, жирів, вуглеводів, вітамінів, мінеральних речовин і води. Продукти розщеплення всіх поживних речовин утворюють у печінці основний метаболічний фон, з якого організм черпає в міру необхідності потрібні для нього речовини. Видалення печінки призводить до загибелі організму.
М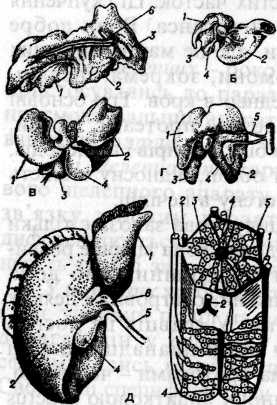 ал.
85. Печінка
хребетних тварин.
ал.
85. Печінка
хребетних тварин.
А - морської черепахи (карети);
Б - летючої миші (нічниці);
В - комахоїдного (їжака);
Г- курки; Д - людини. 1 - ліва частка;
2 - права частка; З - середня частка;
4 — жовчний міхур; 5 - жовчна протока; 6 - печінкова протока. Праворуч - будова часточки печінки людини:
1 —між часточ-кова жовчна протока;
2 - міжчасточкові гілки ворітноївени;
3 - балка; 4 - центральна вена;
5 - венозні капіляри. А, Б, В, Г-вигляд ззаду. Д - вигляд спереду.
Підшлункова залоза
Підшлункова залоза (pancreas) є надзвичайно важливою залозою змішаної секреції. Розвивається вона з ряду невеликих вип'ячувань, що утворюють потім кілька зачатків на початку середньої кишки в тій же ділянці, де закладається й печінка. Найчастіше спостерігається один спинний виріст і два черевні, які пізніше сполучаються разом у спинному мезентерії.
204
Екзокринна частина підшлункової залози має складну альвеолярно-трубчасту будову; вона вкрита тонкою сполучнотканинною капсулою, від якої відходять прошарки сполучної тканини, які ділять паренхіму залози на окремі часточки. Більша частина часток представлена кінцевими секреторними відділами - ацинусами, клітини яких виділяють підшлунковий (панкреатичний) сік. Вивідні протоки часток зливаються в спільні вивідні протоки залози.
Ендокринна частина підшлункової залози представлена особливими групами клітин, розміщеними у вигляді невеликих острівців (скупчень) у товщі залозистих часток. Ці скупчення клітин дістали назву острівців Лангерганса. Вони добре забезпечені кровоносними судинами, але не мають вивідних протоків. Тут виробляються гормони, зокрема, інсулін, глюкагон, які поступають безпосередньо в кров. Три основні групи ферментів підшлункової залози - протеази, ліпази, амілаза - забезпечують розщеплення білків, жирів і вуглеводів, а гормони регулюють вуглеводний обмін, відносну сталість вмісту глюкози в крові, синтез глікогену в печінці.
Серед безхребетних тварин підшлункова залоза є тільки в головоногих молюсків. Серед хордових вона вперше розвивається в безщелепних. У міног, дводишних риб вона розміщена в стінці кишечника; у міксин, осетрових і деяких костистих риб - у тканині печінки. У ссавців це досить компактний орган, що лежить у петлі дванадцятипалої кишки, у яку впадає звичайно двома протоками - черевною, головною (ductus Wirsungi), і спинною, додатковою (ductus Santorini) (мал. 86). У людини підшлункова залоза розміщена в задньочеревному просторі ззаду й нижче шлунка на рівні першого - другого поперекових хребців у вигляді плоского тяжа, витягнутого в горизонтальному напрямку від дванадцятипалої кишки до селезінки. Іннервується підшлункова залоза парасимпатичною і симпатичною нервовими системами.
Отже, у типі хордових можна намітити наступні етапи еволюції травної системи.
У безчерепних зберігається первинний стан кишечної трубки, недиференційованої на окремі органи, крім сліпого печінкового виросту, що функціонує як ворітна система печінки. Характерне пасивне захоплення їжі завдяки рухові
205
війок епітеліальних клітин, що вистилають глоткову ділянку травного тракту.
М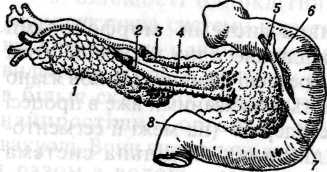 ал.
86. Підшлункова
залоза людини: 1
-
хвіс;
2
- борозенка
селезінкової вени; 3
- борозенка
селезінкової артерії; 4
- тіло;
5
-
головка;
6
-
жовчна
протока; 7
- основна
протока підшлункової залози; 8
- дванадцятипала
кишка.
ал.
86. Підшлункова
залоза людини: 1
-
хвіс;
2
- борозенка
селезінкової вени; 3
- борозенка
селезінкової артерії; 4
- тіло;
5
-
головка;
6
-
жовчна
протока; 7
- основна
протока підшлункової залози; 8
- дванадцятипала
кишка.
У безщелепних - бокової гілки в еволюції хребетних, що пристосувались до паразитичного живлення, утворюється присмоктувальний апарат. Кишечник також не диференційований на відділи.
Усі щелепнороті характеризуються наявністю рухливого щелепного апарату для активного захоплення їжі. У зв'язку з новим характером живлення травна трубка диференціюється на шлунок і різні відділи кишечника, що несуть травні залози (печінка, підшлункова залоза).
У наземних хребетних кишечник досягає значної довжини і має типовий поділ на відділи. Особливої дифференціації досягає він у ссавців, де у зв'язку з характером їжі сформувались різні типи шлунків і відділів кишечника. Своєрідні специфічні риси травної системи в птахів у зв'язку з пристосуванням їх до польоту (відсутність зубів, воло, поділ шлунка на мускульний і залозистий).
207
обміну в акул, морських костистих риб, латимерії, дорослих амфібій та ссавців; сечова кислота - у ящірок, деяких черепах і птахів.
У більшості одноклітинних організмів видільна та репродуктивна системи відсутні. Площа поверхні їх тіла по відношенню до одиниці маси досить велика і тому функцію виділення, а також дихальну, виконує мембрана клітини. Але в більш складних одноклітинних організмів (наприклад, найпростіших) органами виділення служать пульсуючі вакуолі. Вони видаляють зайву рідину з клітинного організму і разом з водою - метаболіти у вигляді СО2, солей та токсичних продуктів обміну. Так вакуолі регулюють водно-сольову рівновагу.
У простих багатоклітинних організмів (плоскі черви, наприклад) з'являється найбільш проста видільна система (протонефридійна), або метаболіти відкладаються в їх тілі в паренхіму, яка виповнює простір між внутрішніми органами. З появою целомічної (вторинної) порожнини тіла метаболіти в розчиненому стані переносяться кровоносною системою через стінку целому в порожнину, а звідти виводяться метанефридійною видільною системою назовні тіла (кільчасті черви, наприклад), або в комах гемолімфою заносяться в міжклітинний простір (міксоцель), а звідти виводяться (шляхом осмосу) мальпігієвими трубочками в кишку і далі - назовні. В інших членистоногих і молюсків існує видозмінена метанефридійна система виділення. І лише в хребетних з'являється типовий нефрон як результат тісного зв'язку кровоносної системи з видільною трубочкою (минаючи порожнини тіла). Поєднання кровоносного клубочка (клубочка Мальпігі) та видільної трубочки через капсулу Боумена і є власне нефроном. Але в хордових в ембріогенезі нирки відтворюють шлях, пройдений ранніми формами хордових. У них утворюється спочатку метанефридійна система (система лійок, які беруть початок із целому, і які потім об'єднуються в один парний вивідний протік назовні). Всередині лійки нефростому є війки, які й виганяють рідину в порожнину трубки з целому. Крім того, поблизу трубки поза целомом утворене скупчення капілярів (так званий тканинний валик - гломус), які також віддають певну кількість фільтрату в цю трубочку. Далі у філогенезі
206