

37. Полиморфизм белков. Типы гемоглобина, лдг и т.Д. Группоспецифические полиморфные системы крови. Полиморфизм белков
У разных особей возникают варианты (мутации) разных генов или варианты одного и того же гена. Варианты генов, образующиеся у отдельных особей, могут постепенно распространяться в популяции в результате наследования, если они не летальны. Так формируется генотипическая неоднородность популяции, которая ведет и к фенотипической неоднородности. На молекулярном уровне фено-типическая неоднородность проявляется как полиморфизм белков — существование разных форм белка, выполняющих одинаковые или очень сходные функции (изобелки). Чаще всего изучают полиморфизм ферментов (т. е. наличие изофер-ментов), поскольку их гораздо легче обнаружить, чем другие белки, по катализируемой ими реакции.
Полиморфизм.
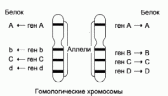
Как известно, место, занимаемое геном в хромосоме (или молекуле ДНК), называют генным локусом. Варианты одного гена в гомологичных хромосомах, занимающие в них гомологичные же локусы (т. е. одинаковое место в одномерном пространстве нитей ДНК), называют аллелями (рис. 5.9). В популяции может быть множество разных аллелей одного гена, в то время как у отдельного индивида — только два, поскольку клетки человека диплоидны (в гаплоидных половых клетках — только один аллель). Гомологичные хромосомы половых клеток в процессе мейоза (профаза I) могут обмениваться аллелями или их частями (рекомбинация). Если при рекомбинации обмениваются не целиком аллельные гены, а участки ДНК меньшей длины, то такой обмен может приводить к появлению новых, прежде не существовавших в популяции аллелей, а не просто новых сочетаний прежних аллелей. После слияния гамет в диплоидном наборе зиготы получаются различные сочетания аллелей, как уже существовавших в популяции, так и вновь возникших в процессе мейоза в паре гамет, образовавших данную зиготу. Рекомбинации — гораздо более частые события, чем мутации, поэтому разнообразие форм внутри вида обусловлено главным образом именно рекомбинациями.Существование в популяции двух или большего числа аллелей одного гена и соответствующих белков называют аллеломорфизмом (частный случай полиморфизма). Аллеломорфизм возникает в процессе филетической эволюции генов. Распространенность (частота) разных аллелей в популяции неодинакова. Принято считать, что аллели с частотой больше 1 % являются полиморфами; если частота меньше, то существование измененного локуса можно объяснить за счет одних только повторных мутаций. При дихотомической эволюции происходит удвоение генов, т. е. образуются новые генные локусы: сначала копии исходного гена, но последующие независимые, неодинаковые мутации копий приводят к появлению в организме изобелков. В этом случае варианты белка являются продуктами разных генных локусов, а не аллельных генов. На практике не всегда легко доказать, являются ли варианты белка продуктами одного локуса, т. е. аллеломорфами, или разных локусов. Таким образом, изобелки — это множественные молекулярные формы белка, обнаруживаемые в пределах организмов одного биологического вида, как результат наличия более чем одного структурного гена в генофонде вида. Множественные гены могут быть представлены как множественные аллели или как множественные генные локусы.
Гемоглобин — это гемопротеин, с молекулярной массой около 60 тыс., окрашивающий эритроцит в красный цвет после связывания молекулы O2 с ионом железа (Fe++). У мужчин в 1 л крови содержится 157 (140—175) г гемоглобина, у женщин — 138 (123—153) г. Молекула гемоглобина состоит из четырех субъединиц гема, связанных с белковой частью молекулы — глобином, сформированной из полипептидных цепей.
Синтез гема протекает в митохондриях эритробластов. Синтез цепей глобина осуществляется на полирибосомах и контролируется генами 11-й и 16-й хромосом. Схема синтеза гемоглобина у человека представлена на рис. 7.2.
ТИПЫ Гемоглобина, содержащий две а- и две В-цепи, называется А-тип (от adult — взрослый). 1 г гемоглобина А-типа связывает 1,34 мл O2. В первые три месяца жизни плода человека в крови содержатся эмбриональные гемоглобины типа Gower I (4 эпсилон цепи) и Gower II (2а и 25 цепи).
Затем формируется гемоглобин F (от faetus — плод). Его глобин представлен двумя цепями а и двумя В. Гемоглобин F обладает на 20—30 % большим сродством к O2, чем гемоглобин А, что способствует лучшему снабжению плода кислородом. При рождении ребенка до 50—80 % гемоглобина у него представлены гемоглобином F и 15—40 % — типом А, а к 3 годам уровень гемоглобина F снижается до 2 %.
Соединение гемоглобина с молекулой кислорода называется оксигемоглобином.
Гемоглобин С- мутантная форма. Эта мутантная форма снижает пластичность эритроцитов организма. В гетерозиготном организме (один аллель, кодирующий нормальный гемоглобин и один мутантный аллель) 28-44 % гемоглобина представлены гемоглобином С, анемия не развивается. У гомозигот почти весь гемоглобин находится в мутантной форме, вызывая умеренную гемолитическую анемию. У таких пациентов кристаллы гемоглобина С можно обнаружить при анализе мазка крови. Присутствие комбинации гемоглобинов С и S вызывает более тяжёлые формы анемии.
Гемоглоби́н S (Hemoglobin S, HbS) — это особая мутантная форма гемоглобина, образующаяся у больных с серповидно-клеточной анемией и склонная к кристаллизации вместо образования нормальной четвертичной структуры и растворения в цитоплазме эритроцита.
Гемоглоби́н A, или ΗbA — нормальный гемоглобин взрослого человека.
Этот белок представляет собой тетрамер, состоящий из двух пар полипептидных цепей — мономеров: двух мономеров α-цепей и двух мономеров β-цепей (так называемый гемоглобин A, или гемоглобин α2β2), или двух мономеров α и двух мономеров δ (гемоглобин Α2, или гемоглобин α2δ2).
Гемоглобин A — HbA (α2β2)
Гемоглобин A2 — HbA2 (α2δ2)
В эритроцитах здорового взрослого человека гемоглобин А (α2β2) является основным вариантом гемоглобина и составляет в норме почти 97 % общего гемоглобина эритроцитов. Оставшиеся около 3 % приходятся на гемоглобин Α2 (α2δ2). Количество этой формы гемоглобина увеличивается у больных β-талассемией.
Гемоглобин A (англ. Adult — взрослый) относится к сложным белкам — хромопротеидам. Он состоит из белковой и небелковой частей. Белковая часть представлена четырьмя полипептидными цепями, которые попарно одинаковы: 2α и 2β цепи. В связи с тем, что цепи гемоглобина A неодинаковы, он относится к гетеротетрамерным молекулам. Каждая полипептидная цепь связана с небелковой частью, которую представляет гем. Комплекс гема с α- или β-полипептидной цепью представляет субъединицу или протомер. Таким образом, вся молекула гемоглобина состоит из 4-х субъединиц или протомеров, образуя олигомер.
В структуре HbA находится 574 аминокислоты. Полипептидные цепи гемоглобина на 80 % спирализованы, обозначаются спирали буквами латинского алфавита от A до H. Неспиральные участки обозначаются двумя буквами латинского алфавита, между спиралями которых они находятся. Это необходимо для обозначения молекулярных замещений, приводящих к синтезу аномальных гемоглобинов.
ЛДГ
Лактатдегидрогеназа (L-лактат: NAD-оксидоредуктаза (LDH) 1.1.1.27) — фермент, принимающий участие в реакциях гликолиза. Лактатдегидрогеназа катализирует превращение лактата в пируват, при этом образуется HADH+.
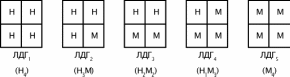
Молекула любого изофермента ЛДГ распадается при обработке мочевиной на 4 субъединицы. Субъединицы представляют собой полипептиды двух основных типов — Н и М, различающихся по аминокислотному составу, иммунохимическим свойствам.
Изоферменты ЛДГ представляют собой тетрамеры из субъединиц двух типов в каждой молекуле изофермента, при этом число субъединиц равно 4. Это может быть представлено в виде следующей схемы:
 Н
и М это субъединицы фермента.
ЛДГ1
– Н4 – только в миокарде;
Н
и М это субъединицы фермента.
ЛДГ1
– Н4 – только в миокарде;
ЛДГ2 – Н3М1 – в миокарде, эритроцитах, др.;
ЛДГ3 – Н2М2 – в легких;
ЛДГ4 – Н1М3 – в скелетных мышцах, печени;
ЛДГ5 – М4 – в печени.
Группоспецифические полиморфные системы крови я найти не смог.