

28 Неструктурированные модели клеточного роста и образования продуктов жизнедеятельности.
В простом случае образование метаболитов, утилизация субстрата и клеточный рост связаны простым стехиометрическим уравнением. Скорость образования продуктов метаболизма выражается:
rfp= -Ip/s∙rfs (1)
rfp= -Ip/x∙rfx (2)
Пример спиртовое брожение. Такого типа кинетика образования продуктов жизнедеятельности является сопряженной с ростом клеток.
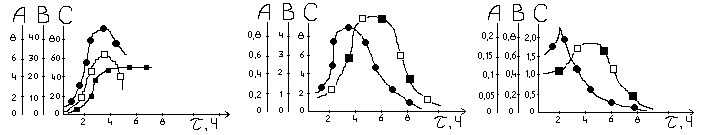
(а) (б) (в)
 масса сухого
мицелия, г/л; рост клеток, г/л∙ч;
рост клеток, г/г∙ч;
масса сухого
мицелия, г/л; рост клеток, г/л∙ч;
рост клеток, г/г∙ч;
 синтез спирта,
г/л; синтез спирта, г/л;
синтез спирта, г/г∙ч;
синтез спирта,
г/л; синтез спирта, г/л;
синтез спирта, г/г∙ч;
 утилизация
сахара, г/л; утилизация сахара,
г/л∙ч; утилиз. сахара, г/г∙ч
утилизация
сахара, г/л; утилизация сахара,
г/л∙ч; утилиз. сахара, г/г∙ч
τ – время процесса брожения, ч
Для спиртового брожения характерна простая, сопряженная с ростом кинетика образования продуктов метаболизма. На рисунках указано изменение во времени массы образующегося спирта, утилизация субстрата (а). Изменение во времени объемной и удельной скорости образования биомассы, утилизация субстрата и синтеза продукта метаболизма (спирта) (б,в). Если речь идет о вторичных метаболитах, на первой стадии периодического процесса продукты жизнедеятельности не образуются, вплоть до начала стационарной фазы или позже. В этих случаях кинетика образования продуктов жизнедеятельности удовлетворяет описанной простой не сопряженной с ростом модели, в которой скорость образования продуктов метаболизма принимается пропорциональной не скорости роста клеток, а их концентрации.
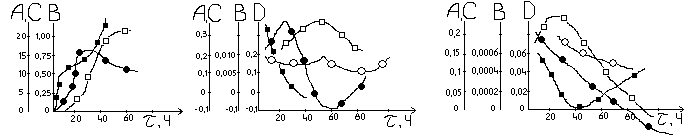
(г) (д) (е)
масса сухого мицелия, г/л; клеточный рост, г/л∙ч; клет. рост, г/г∙ч;
синтез пенициллина, г/л; синтез пенициллина, г/л∙ч; синтез пениц., г/г∙ч;
утилизация сахара, г/л; утилизация сахара, г/л∙ч; утилиз. сах., г/г∙ч;
τ –
время ферментации, ч
поглощение кислорода, г/л∙ч поглощ.
кисл., г/г∙ч;
–
время ферментации, ч
поглощение кислорода, г/л∙ч поглощ.
кисл., г/г∙ч;
В классической работе Льюдикина-Пайрета по изучению молочнокислых бактерий Lact. delbrueckii показано, что в кинетику образования продуктов метаболизма делают вклад как сопряженные с ростом факторы, так и несопряженные.
rfp=αrfx+βx (1)
Уравнение (1) позволяет интерпретировать экспериментальные результаты многих микробиологических процессов. Такая форма зависимости возникает тогда, когда изучаемое вещество является конечным продуктом метаболитического пути, связанного с выделением энергии, например, в некоторых процессах анаэробного брожения. В этих случаях первое и второе слагаемые в уравнениях рассматривают как меру энергии, расходуемой на клеточный рост и на поддержание клетки соответственно.
29 Структур-е модели клет-о роста Компарт-ые модели (Уильямса, Хардера – Роуэлса). Метаболич-е модели клет-о роста. Структур-е модели образ-я прод-в жизнедеят-и. Хим-и структур-е модели кинетики образования продуктов метаболизма клеток. Морфологически структурированная модель кинетики биосинтеза ЦСС.
Метаболические модели клеточного роста. Рассмотрим метаболическую модель Бийкерга и Холла. подразумевается число молей или единиц активности для фермента).
1. брожение: (ra) Aw+a1+Sw+Ej → 2 Bw+a2Ew + Ej+CO2
2. дыхание: (rв) Aw+a3 Ew+ O2+ Er+Ef → 2Bw+Er+Ef+CO2
3. деление: Bw → Aw, где ra, rв, rc - выражаются следующими формулами:
rA=![]()
![]() ;
rB=
;
rB=
![]() ;
rC=K∙b
;
rC=K∙b
контрольные
концентрации ферментов: eR
=![]() ;
eF
=
;
eF
=![]()
Скор-и биосинтеза
ферме-ых сис-м брожения и дых-я будут
иметь следующие выраж-я: (rF)E
брож.=
rA+r5Vc+k4(eF
–eF);
(rF)E
дых.= rB
+k6
(eR
–er)
В этих ур-х er
и ef
обоз-ют активности ферм-в в единице
объема культуры В основу структурированных
моделей
клеточного
роста
закладывают приближения и допущения
при кот. эта модель раб-т и при кот-й
расчетные данные мах полно совпадают
с экспер-ми данными. В структур-е модели
в кач-ве переем-й могут прим-ся масс-я
доля xj,
моляр-я доля cj
дан-о биол-го
объекта в един-е объема. При усл-и пол-го
перемеш-я урав-е матер-го баланса по
компоненту j:
![]() VR
x
rAj
+
VR
x
rAj
+![]() Фк
∙ сj,
(1) Если допустить, что ρс
и Vρ
не изменяются во времени, то после
диффер-я урав-я (1) получим
Фк
∙ сj,
(1) Если допустить, что ρс
и Vρ
не изменяются во времени, то после
диффер-я урав-я (1) получим
![]() rfi
– Cj
rfi
– Cj
![]() +
Cj
+
Cj
![]() (2), для период-х реак-в Фх=
0, след-но,
rfj
– μ Cj
(3). В стерил-х услов-х приведении питат-х
стерил-х в-в в ПРПП, оценка правой части
уравнения (2) приводит к ур-ю (3). В простейшей
структур-й модели предусматр-ся
компартментация
биомассы в небольшом кол-е компон-в.
Уильямс
предложил двухкомпартмент-ю модель,
кот-я достаточно точно описывает нек-е
стороны динамики роста в период-м
процессе 4)Биомасса построена только
из компонентов (1) и (2)
(2), для период-х реак-в Фх=
0, след-но,
rfj
– μ Cj
(3). В стерил-х услов-х приведении питат-х
стерил-х в-в в ПРПП, оценка правой части
уравнения (2) приводит к ур-ю (3). В простейшей
структур-й модели предусматр-ся
компартментация
биомассы в небольшом кол-е компон-в.
Уильямс
предложил двухкомпартмент-ю модель,
кот-я достаточно точно описывает нек-е
стороны динамики роста в период-м
процессе 4)Биомасса построена только
из компонентов (1) и (2)
![]() k1sx
(4);
k1sx
(4);
![]() k1sx
(5);
k1sx
(5);
![]() k1s(ρ1+ρ2)-k2ρ1ρ2-ρ1μ
(6)
k1s(ρ1+ρ2)-k2ρ1ρ2-ρ1μ
(6)
![]()
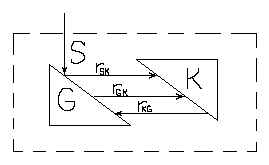 k2
ρ1ρ2-
μρ2
(7); μ=k1s (8);
(ρ1+ρ2)=
ρC=
const (9)
k2
ρ1ρ2-
μρ2
(7); μ=k1s (8);
(ρ1+ρ2)=
ρC=
const (9)
тогда уравнение
(3) будет записано следующим образом:
![]() -k1sf2+(k2
ρC)
f2(1-f2)
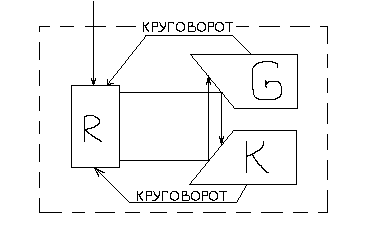
Двухкомпартментальная
модель Хардера и Роуэлса
В этой простой двухкомпартментальной
модели взаимодействие G
и К осуществляется путем полимеризации
и деполимеризации, что соответствует
состоянию метаболизма поддержания. В
случае трехкомпартментальной модели
Хардер и Роуэлс предусмотрели более
четкую определенную биохимическую
природу компонентов. К – обозначает
РНК; G
– белок; R
– другие компоненты биомассы. Метаболизм
поддержания отражен как кругооборот
компонентов К и G.
-k1sf2+(k2
ρC)
f2(1-f2)
Двухкомпартментальная
модель Хардера и Роуэлса
В этой простой двухкомпартментальной
модели взаимодействие G
и К осуществляется путем полимеризации
и деполимеризации, что соответствует
состоянию метаболизма поддержания. В
случае трехкомпартментальной модели
Хардер и Роуэлс предусмотрели более
четкую определенную биохимическую
природу компонентов. К – обозначает
РНК; G
– белок; R
– другие компоненты биомассы. Метаболизм
поддержания отражен как кругооборот
компонентов К и G.
Механизмы белкового синтеза клетки имеют большое значение для производства ферментов, гормонов, полипептидов. В случае расчетов по химически структурированным моделям в допущения предполагается оптимальный уровень начальной концентрации фосфата, кот. снижается по мере расходования его клеткой. Рассмотрим модель биосинтеза алкалоидов культурой клеток Cloviseps purpurea, учитывается клеточный рост периодической культуры с образованием продукта жизнедеятельности – алкалоида.
Рост клеток:
 ;
2. концентрация фосфатов в среде:
;
2. концентрация фосфатов в среде:
 ;
3. концентрация фосфатов в клетке:
;
3. концентрация фосфатов в клетке:
 ;
4. образование алкалоидов:
;
4. образование алкалоидов:
 .
.
Выражение k2x2 отражает скорость лизиса клеток.Первое слагаемое уравнения 2 описывает активный транспорт фосфата в клетку.
ЦСС: исследования проводились на примере глубинной культуры Cephalosporium acremonium. Существует 3 различные морфологические формы: гифы (h), набухшие
фрагменты гиф (s) и артроспоры (а). В зависимости от среды эти формы могут взаимопревращаться: набухшие фрагменты
г
 ифы
гиф артроспоры
ифы
гиф артроспоры


 а
а
h s истощение глюкозы

 ассимиляция
индукция эндогенным метионином
ассимиляция
индукция эндогенным метионином
метионина и катабалитная репрессия глюкозы
 глюкозы
глюкозы
ферменты, синтезирующие ЦСС ЦСС
ЦСС продуцируется набухшими фрагментами гиф. Скорость синтеза индуцируется внутриклеточным метионином и подавляется глюкозой.
1.рост и диф-ция
плесени:![]() ;
;
![]() ;
;
![]() ,
где β(g,m)=
,
где β(g,m)=![]() ,
,
![]() .
Общая биомасса разделена на 3 типа.
Концентрация глюкозы – g,
а метионина – m. Модель
утилизации субстрата подразумевает,
что g и m
усваиваются только формами h
и s.
.
Общая биомасса разделена на 3 типа.
Концентрация глюкозы – g,
а метионина – m. Модель
утилизации субстрата подразумевает,
что g и m
усваиваются только формами h
и s.
2.Утилизация глюкозы и метионина:
 ;
;
![]() ,
где
,
где
![]() ,
,
![]() .
Средняя внутриклеточная концентрация:
.
Средняя внутриклеточная концентрация:
![]() ,
,
![]() .
Такая детальная модель описывает все
стороны процесса. Сравнением найденных
и расчетных величин
.
Такая детальная модель описывает все
стороны процесса. Сравнением найденных
и расчетных величин
![]() определяют основные параметры в
уравнениях модели.3) Синтез ферменов:
определяют основные параметры в
уравнениях модели.3) Синтез ферменов:![]() ,
,
 .
4) Образование антибиотика:
.
4) Образование антибиотика:
![]() .
.