

80. Почему у дрозофилы генотип хо приводит к развитию мужского пола, хху- женского, а у человека – наоборот?
Хотя у дрозофилы и у человека гетерогаметный пол мужской, и тип определения пола ХУ, т.е. соматичесие клетки обычно женского организма имеют две ХХ, хромосомы, а мужского – ХУ, но все же механизмы хромосомного определения пола не одинаковы у этих видов. У человека У-хромосома является обязательной для определения пола, и в случае анеуплоидии при любом количестве Х хромосом, У-хромосома обуславливает развитие мужского пола. У дрозофилы роль У-хромосомы не является определяющей, для развития мыжского пола. Разитие женского или мужского пола в этом случае от соотношения количества Х-хромосом и количества наборов аутосом(А). При генотипе ХО, соотношение nX/nA=1/2, что соответствует мужскому полу. При генотипе ХХУ, соотношенние nX/nA=2/2=1, что соответствует мужскому полу.
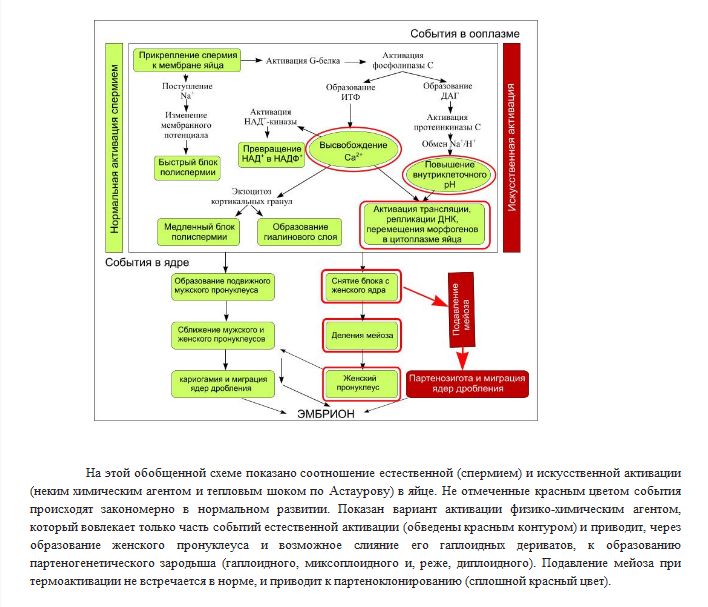
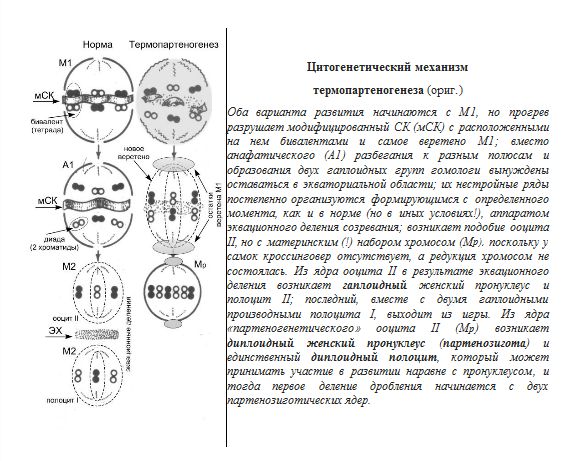
81.Искусственный партеногенез и его цитологические механизмы
Искусственный П. у животных был впервые получен русским зоологом А. А. Тихомировым. Он показал (1886), что неоплодотворённые яйца тутового шелкопряда можно побудить к развитию растворами сильных кислот, трением и др. физико-химическими раздражителями. В дальнейшем искусственный П. был получен Ж. Лёбом и др. учёными у многих животных, главным образом у морских беспозвоночных (морские ежи и звёзды, черви, моллюски), а также у некоторых земноводных (лягушка) и даже млекопитающих (кролик). В конце 19 — начале 20 вв. опыты по искусственному П. привлекали особое внимание биологов, давая надежду с помощью этой физико-химической модели активации яйца проникнуть в сущность процессов оплодотворения. Искусственный П. вызывают действием на яйца гипертонических или гипотонических растворов (так называемый осмотический П.), уколом яйца иглой, смоченной гемолимфой (так называемый травматический П. земноводных), резким охлаждением и особенно нагревом (так называемый температурный П.), а также действием кислот, щелочей и т.п. С помощью искусственного П.обычно удаётся получать лишь начальные стадии развития организма; полный П. достигается редко, хотя известны случаи полного П. даже у позвоночных животных (лягушка, кролик). Способ массового получения полного П., разработанный (1936) для тутового шелкопряда Б. Л. Астауровым, основан на точно дозированном кратковременном прогреве (до 46 °С в течение 18 мин)извлечённых из самки неоплодотворённых яиц. Этот способ даёт возможность получать у тутового шелкопряда особи только женского пола, наследственно идентичные с исходной самкой и между собой. Получаемые при этом ди-, три- и тетраплоидные клоны можно размножать посредством П. неограниченно долго. При этом они сохраняют исходную гетерозиготность и "гибридную силу". Отбором получены клоны, размножающиеся посредством П. так же легко, как обоеполые породы посредством оплодотворения (более 90% вылупления активированных яиц и до 98% жизнеспособности). П. представляет разносторонний интерес для практики шелководства.
82.Исскуственный синтез генов

+



83.Явление гетерозиса у человека, проявление и значение
При отдаленном скрещивании можно наблюдать появление гибридов с повышенной жизнеспособностью в первом поколений. Это явление получило название гетерозиса. Причиной гетерозиса является перевод вредных рецессивных мутаций в гетерозиготное состояние, при котором они не проявляются в фенотипе.
Эффект гетерозиса во всех учебниках описывается на примере выведения новых сортов кукурузы. Смысл сводится к тому, что если сначала вывести чистые сорта, а потом их скрещивать, качество дочерних растений будет значительно выше родительского. В первом поколении эффект максимален, во втором - слабее и т.д. до полного вырождения. Гетерозисная кукуруза плохо воспроизводится сама. Рано или поздно гетерозисные линии вырождаются окончательно и погибают. Потому для выращивания гетерозисной кукурузы семена должны производить отдельные специализированные на чистых линиях хозяйства.
Для гетерозиса можно только выдвигать гипотезы, например: взаимодействие двух сильно различающихся генов в одном комплекте может давать третье свойство - положительное. Генный компект работает в паре. При образование новых хромосом участок кода взаимодействия делится неправильно, и половина суммы положительного кода - это дефектный код. К примеру, существуют условные генные компоненты 15 и 25. Если они присутствуют у человека, это дает владельцу положительные качества. Но при наследовании получаются соотношения 22 и 18 – что дает качества отрицательные.
Другая версия: при гетерозисе в комплектах оказываются максимально разные хромосомы. Гетерозис гасит рецессивные проявления. Разность между хромосомами обуславливает высокую степень перекрещивания белковых цепочек. Потому в науке и существует мнение о пониженной жизнеспособности смешанных типов. Но у человека дети с разрушительными явлениями перекрещивания обычно не рождаются, как и у всех сложных существ. Поскольку не родившиеся (микровыкидыши) не учитываются, то и создается иллюзия повышенной жизнеспособности.
В науке существует, но не популяризируется мнение, что межнациональные дети имеют более низкое биологическое качество. Это мнение преподносится, но никогда не доказывается. На самом деле это не совсем так – гетерозис приводит к тому, что в первом поколении дети имеют качество выше родителей. Но кто-то сделал намек. Качество падает в последующих поколениях. Но доказывать эти сведения никто не стал – скорее всего, по этическим соображениям не захотел разбираться с подобными проблемами.
В результате смешанных браков первое поколение имеет качество выше родительского, а со второго может наступить резкий спад до полного вырождения. Явление гетерозиса, возможно, преодолимо подбором национальных пар для каждого поколения с возвратом в нацию. Но над этим вопросом никто не работает, и на создание методики могут потребоваться десятилетия. Лица с эффектом гетерозиса не могут быть отнесены к какой-либо нации; но в некоторых случаях их потомки могут быть отнесены к нациям (биологический эффект расщепления). Действительно, у родителей, произошедших от смешанных браков, и потому не относящихся к национальностям, могут рождаться дети разных национальностей.
При межнациональном скрещивании территориальные генетические преимущества нивелируются, а национальные генетические заболевания вырываются из локальных «ящиков Пандоры» во внешний мир, постоянно умножаясь.
Люди чисто национальные живут в среде межнационалного смешивания. Нация, как яичный желток, плавает в гетерозисном белке, но в отличие от подобной схемы, четкой цветовой границы у нее не прослеживается. Прослеживаются состояния – или эффект гетерозиса выражен, или он отсутствует. Или человек – представитель чистой национальной линии, или он не является чистым национальным экземпляром.
Существует еще одна особенность: отдаленное скрещивание называется аутбридингом – при этом варианте качество почти незаметно, но возрастает. Разница между гетерозисом и аутбридингом пока что не может быть точно определена. Остается предположить, что при очень близких национальных вариантах, имеющих один ландшафт исходным, получается эффект аутбридинга. При более отдаленных - гетерозиса. А где кончается «более» и начинается «менее» - неизвестно.
Существуют нации родительские и дочерние. Русские - пример давнего смешивания, результат древнего массового гетерозиса славян и угро-финнов. Но сейчас есть еще и несмешанные славяне - белорусы, и несмешанные угро-финны - карелы. Так что в результате смешивания русских с этими двумя нациями получаются русские безо всякого гетерозисного эффекта. А при смешивании белорусов и угро-финнов рост качества в слабой степени присутствует, но он не приводит к вырождению линии. Выходит, что эффекта гетерозиса нет, а есть эффект аутбридинга. Русские и есть результат этого смешивания. Скорее всего, это связано с формированием обеих наций в почти одинаковом ландшафте.
Для родителей гетерозис несет эффект отрицательный, увеличивая отбраковку потомства. Но он увеличивает возможность появления единичных экземпляров, более чем аборигены адаптированных к природной среде – опять-таки с шансом передачи генов по наследству. Положительный признак через десятки поколений становится национальным.
Гетерозис может давать положительный эффект только при очень высокой смертности – свыше 50%, принятых как норма для чистых наций. При смешанном браке для гетерозисных людей степень отбраковки неизвестна, возможно, варьируется в зависимости от степени разброса параметров, но скорее всего должна составлять свыше 75%. При идеальном гетерозисном браке, т.е при первобытном здоровье родителей, большинство детей все равно родятся не адаптированными к природной среде проживания.
При гетерозисе появляются особи, имеющие не узкоспециализированные свойства адаптации, а универсальную адаптацию. Т.е. утрачиваются признаки адаптации к ландшафту, но вместо них могут появиться высокая степень интеллекта, большая физическая сила, очень часто появляются очень хорошие способности к изучению иностранных языков. Универсальность первого гетерозисного поколения дает ему преимущества при жизни в искусственных ландшафтах, например, в городах. Или на вновь заселенных территориях, в условиях стран - искусственных ландшафтов. Возможно, это ненаучно, но люди с эффектом гетерозиса превосходят по средним качествам чистые национальные экземпляры с тем, что чтобы дать большее потомство для обеспечения последующей большей отбраковки.
Люди с эффектом гетерозиса очень часто оказываются действительно талантливыми людьми, которых не понимают окружающие. А из окружающих их чаще всего понимают люди со сходным уровнем гетерозисного эффекта.
Гетерозис является одним из компонентов генетического дрейфа – с тем, как изменяются условия, нация должна изменять свой генетический код. Но нация не знает, в какую сторону она «должна» «дрейфовать». Люди с гетерозисным эффектом являются переносчиками успешных генов из нации в нацию. Но поскольку неизвестно, какие из этих генов нации понадобятся, а какие нет, то интеграция в нацию требует большого числа положительных случайностей.
Не каждый гетерозисный экземпляр может интегрироваться в нацию, и далеко не каждый может привнести в нее положительный момент. Гетерозисные предки были у каждого человека, но у большинства смешанных браков не осталось потомков.
Гетерозис – это необходимый элемент человеческого развития, но он требует соблюдения двух условий – повышенная отбраковка потомства и хотя бы видимое отсутствие генетических проблем у родителей, вступающих в межнациональный брак. Гетерозис должен присутствовать в гомеопатических дозах и только для близких генетически популяций.
Гетерозис только тогда приводит к появлению новых наций, когда имеет место глобальное смешение исходного материала с высокой отбраковкой. А при небольших в процентном отношении вариантах смешивания он дает нации большую вариабельность признаков адаптации.
Варианты межнационального смешивания можно классифицировать следующим образом:
При смешивании близких одноландшафтных наций эффект практически не проявляется, соответственно, и серьезного качества впоследствии не наблюдается. Но нет и серьезного падения. Если нет ни серьезного роста параметров, ни их отката, то это не эффект гетерозиса, а эффект аутбридинга.
При смешивании внутри локальных рас и дальних наций одной расы наблюдается достаточный, чтобы заметить, эффект, особенно в интеллектуальной сфере. Далее – если линия продолжается в доминантной расе, то негативный эффект минимален; если в рецессивной – качество начинает падать из поколения в поколение.
При смешивании основных рас может наблюдаться как небольшой положительный эффект в интеллекте и физическом состоянии, так и отрицательный. Если линия с положительным эффектом продолжается в доминантной расе, эффект фактически отсутствует. Если такая линия продолжается в рецессивной расе, эффект приводит к вырождению линии. Линия с первичным отрицательным эффектом обречена на вырождение.
Вторичный гетерозис – смешивание людей с гетерозисным эффектом – не приводит к повышению биологического качества и обычно всегда ведет линию к вырождению.
По гетерозисному эффекту можно сделать следующие выводы:
В стабильно существующей популяции гетерозис необходим, но в гомеопатических, в очень малых дозах. Природа дальше сама разберется, нужны популяции эти экземпляры или нет.
При смешивании целых наций получаются новые нации. Результат может быть положительным, но чаще оказывается отрицательным.
В зависимости от силы эффекта гетерозиса его можно повторять через несколько поколений.
За все нужно платить, и за гетерозис - в частности.
Великие успехи современной цивилизации, и особенно Америки в технической сфере, имеют одну причину - это эффект гетерозиса в результате межнационального смешивания. Дети от межнациональных браков интеллектуальнее; но внуки резко деградируют. В силу незнания, что гетерозисные линии вырождаются, оказалось очень много людей, принесших свою линию преемственности поколений в жертву интеллектуальному развитию цивилизации. Но потенциал гетерозиса должен подойти к концу, и тогда - сначала количественная катастрофа с гибелью большей части гетерозисных линий, потом формирование новых чистых линий и далее медленное и постепенное развитие.
Сцепленное наследование. У. Сэттон и Р. Пеннет в 1908 г. обна-ружили отклонения от свободного комбинирования признаков согласно третьему закону Менделя. В 1911-12 г.г. Т. Морган и сотр. описали явление сцепления генов - совместную передачу группы генов из поколения в поколение. Опыты проводились на мухах дрозофилах с учетом двух пар альтернативных признаков - серый и черный цвет тела, нормальные и короткие крылья. При скрещивании гомозиготных особей с серым телом и нормаль-ными крыльями с особями с черным телом и короткими крыльями получено единообразие гибридов первого поколения, особи которого имели доминантные признаки:
 Для
выяснения генотипа гибридов I поколения
Морган провел анализирующее скрещивание.
Он взял рецессивную гомозиготную самку
и скрестил ее с дигетерозиготным самцом,
затем провел реципрокное скрещивание:
Для
выяснения генотипа гибридов I поколения
Морган провел анализирующее скрещивание.
Он взял рецессивную гомозиготную самку
и скрестил ее с дигетерозиготным самцом,
затем провел реципрокное скрещивание:
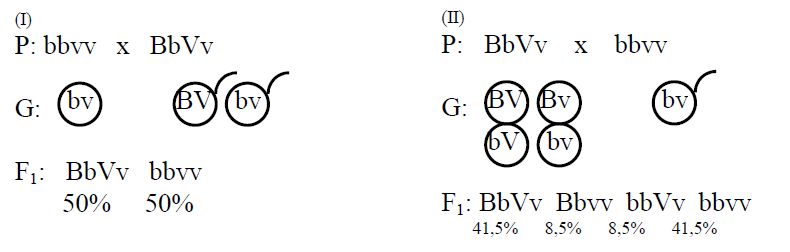
При свободном комбинировании генов согласно третьего закона Менделя в поколении должны были появиться мухи четырех разных фенотипов по 25%. Это можно пояснить следующей схемой:
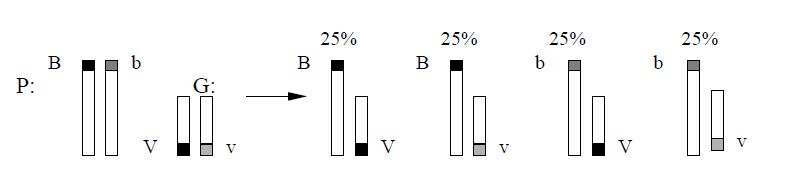
Но гены разных аллельных пар могут находиться в одной паре гомологичных хромосом.
Одна из пары гомологичных хромосом содержит 2 доминантных гена (BV), а другая - 2 рецессивных (bv). В процессе мейоза одна хромосома с генами BV попадет в одну гамету, а другая (с генами bv) - в другую. Таким образом, у дигетерозиготного организма образуется не четыре, а только два типа гамет и потомки будут иметь такое же сочетание признаков, как и родители. В данном случае сцепление будет полным.
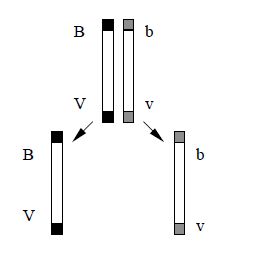
При изучении результатов второго скрещивания (II) было обнаружено нарушение полного сцепления генов. Если взять дигетерозиготную самку мухи дрозофилы и скрестить ее с рецессив-ным самцом, то получается 4 разновидности фенотипов потомков: 41,5% с серым телом и длинными крыльями, 41,5% с черным телом и короткими крыльями и по 8,5% гибридных форм - с серым телом и короткими крыльями и с черным телом и длинными крыльями. В этом случае сцепление оказывается неполным, т. е. происходит перекомбинация генов, локализованных в одной хромосоме.
Это обьясянется кроссинговером.
Кроссинговер
Кроссинговер (от англ. crossing–over – перекрёст) – это обмен гомологичными участками гомологичных хромосом (хроматид).
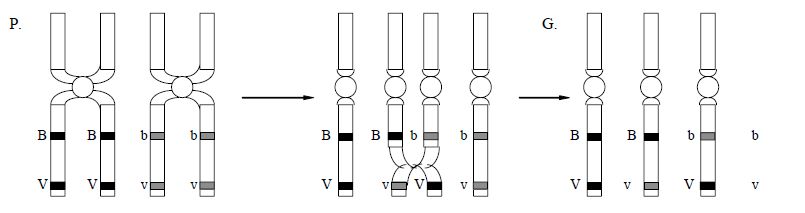
Цитогенетика кроссинговера
Механизм кроссинговера «разрыв–воссоединение»
Согласно теории Янссенса–Дарлингтона, кроссинговер происходит в профазе мейоза. Гомологичные хромосомы с гаплотипами хроматид АВ и ab образуют биваленты. В одной из хроматид в первой хромосоме происходит разрыв на участке А–В, тогда в прилежащей хроматиде второй хромосомы происходит разрыв на участке a–b. Клетка стремится исправить повреждение с помощью ферментов репарации–рекомбинации и присоединить фрагменты хроматид. Однако при этом возможно присоединение крест–накрест (кроссинговер), и образуются рекомбинантные гаплотипы (хроматиды) Ab и аВ. В анафазе первого деления мейоза происходит расхождение двухроматидных хромосом, а во втором делении – расхождение хроматид (однохроматидных хромосом). Хроматиды, которые не участвовали в кроссинговере, сохраняют исходные сочетания аллелей. Такие хроматиды (однохроматидные хромосомы) называются некроссоверными; с их участием разовьются некроссоверные гаметы, зиготы и особи. Рекомбинантные хроматиды, которые образовались в ходе кроссинговера, несут новые сочетания аллелей. Такие хроматиды (однохроматидные хромосомы) называются кроссоверными, с их участием разовьются кроссоверные гаметы, зиготы и особи.
Таким образом, вследствие кроссинговера происходит рекомбинация – появление новых сочетаний (гаплотипов) наследственных задатков в хромосомах.
Интерференция – это подавление кроссинговера на участках, непосредственно прилегающих к точке происшедшего обмена. Рассмотрим пример, описанный в одной из ранних работ Моргана. Он исследовал частоту кроссинговера между генами w (white – белые глаза), у (yellow – желтое тело) и m (miniature – маленькие крылья), локализованными в Х-хромосоме D. melanogaster. Расстояние между генами w и у в процентах кроссинговера составило 1,3, а между генами у и m – 32,6. Если два акта кроссинговера наблюдаются случайно, то ожидаемая частота двойного кроссинговера должна быть равна произведению частот кроссинговера между генами у и w и генами w и m. Другими словами, частота двойных кроссинговеров будет 0,43%. В действительности в опыте был обнаружен лишь один двойной кроссинговер на 2205 мух, т. е. 0,045%. Ученик Моргана Г. Меллер предложил определять интенсивность интерференции количественно, путем деления фактически наблюдаемой частоты двойного кроссинговера на теоретически ожидаемую (при отсутствии интерференции) частоту. Он назвал этот показатель коэффициентом коинциденции, т. е. совпадения. Меллер показал, что в Х-хромосоме дрозофилы интерференция особенно велика на небольших расстояниях; с увеличением интервала между генами интенсивность ее уменьшается и на расстоянии около 40 морганид и более коэффициент коинциденции достигает 1 (максимального своего значения).
Двойной и множественный кроссинговер
Т. Морган предположил, что кроссинговер между двумя генами может происходить не только в одной, но и в двух и даже большем числе точек. Четное число перекрестов между двумя генами, в конечном счете, не приводит к их перемещению из одной гомологичной хромосомы в другую, поэтому число кроссинговеров и, следовательно, расстояние между этими генами, определенное в эксперименте, снижаются. Обычно это относится к достаточно далеко расположенным друг от друга генам. Естественно, что вероятность двойного перекреста всегда меньше вероятности одинарного. В принципе она будет равна произведению вероятности двух единичных актов рекомбинации. Например, если одиночный перекрест будет происходить с частотой 0,2, то двойной – с частотой 0,2 × 0,2 = 0,04. В дальнейшем, наряду с двойным кроссинговером, было открыто и явление множественного кроссинговера: гомологичные хроматиды могут обмениваться участками в трех, четырех и более точках.
Цитологическое (цитогенетическое) доказательство кроссинговера
Прямые цитологические свидетельства обмена частей хромосом во время кроссинговера были получены в начале 30-х годов у дрозофилы и кукурузы. Рассмотрим опыт Штерна, проведенный на D. melanogaster. Обычно две гомологичные хромосомы морфологически неразличимы. Штерн исследовал Х-хромосомы, которые имели морфологические различия и, следовательно, были гомологичны не полностью. Однако гомология между этими хромосомами сохранялась на большей части их длины, что позволяло им нормально спариваться и сегрегировать в мейозе (то есть нормально распределяться по дочерним клеткам). Одна из Х-хромосом самки в результате транслокации, т. е. перемещения фрагмента Y-хромосомы, приобрела Г-образную форму. Вторая Х-хромосома была короче нормальной, так как часть ее была перенесена на IV хромосому. Были получены самки, гетерозиготные по указанным двум, морфологически различным, Х-хромосомам, а также гетерозиготные по двум генам, локализованным в Х-хромосоме: Bar (В) и carnation (cr). Ген Bar – это полудоминантный ген, влияющий на количество фасеток и, следовательно, форму глаза (мутанты с аллелем В имеют полосковидные глаза). Ген cr контролирует окраску глаз (аллель cr+ обусловливает нормальную окраску глаз, а аллель cr – окраску глаз цвета красной гвоздики). Г-образная Х-хромосома несла аллели дикого типа В+ и cr+, укороченная хромосома – мутантные аллели В и cr. Самки указанного генотипа скрещивались с самцами, имевшими морфологически нормальную Х-хромосому с аллелями cr и В+. В потомстве самок было два класса мух с некроссоверными хромосомами (crB / crB+ и cr+B+ / crB+) и два класса мух, фенотип которых соответствовал кроссоверам (crB+ / crB+ и cr+B / crB+). Цитологическое исследование показало, что у кроссоверных особей произошел обмен участками Х-хромосом, и, соответственно, изменилась их форма. Все четыре класса самок имели по одной нормальной, т. е. палочковидной, хромосоме, полученной от отца. Кроссоверные самки содержали в своем кариотипе преобразованные в результате кроссинговера Х-хромосомы – длинную палочковидную или двуплечую с короткими плечами. Эти опыты, так же как и одновременно полученные аналогичные результаты на кукурузе, подтвердили гипотезу Моргана и его сотрудников о том, что кроссинговер представляет собой обмен участками гомологичных хромосом и что гены действительно локализованы в хромосомах.
Соматический (митотический) кроссинговер
В соматических клетках иногда происходят обмены между хроматидами гомологичных хромосом, в результате которых наблюдается комбинативная изменчивость, подобная той, которая регулярно генерируется мейозом. Нередко, особенно у дрозофилы и низших эукариот, гомологичные хромосомы синаптируют в митозе. Одна из аутосомно-рецессивных мутаций человека, в гомозиготном состоянии приводящая к тяжелому заболеванию, известному под названием синдром Блюма, сопровождается цитологической картиной, напоминающей синапсис гомологов и даже образование хиазм. Доказательство митотического кроссинговера было получено на дрозофиле при анализе изменчивости признаков, определяемых генами у (yellow – желтое тело) и sn (singed – опаленные щетинки), которые находятся в Х-хромосоме. Самка с генотипом ysn+ / y+sn гетерозиготна по генам у и sn, и поэтому в отсутствие митотического кроссинговера ее фенотип будет нормальным. Однако если кроссинговер произошел на стадии четырех хроматид между хроматидами разных гомологов (но не между сестринскими хроматидами), причем место обмена находится между геном sn и центромерой, то образуются клетки с генотипами y sn+ / y+ sn+ и y+sn/y+n. В этом случае на сером теле мухи с нормальными щетинками появятся близнецовые мозаичные пятна, одно из которых будет желтого цвета с нормальными щетинками, а другое – серого цвета с опаленными щетинками. Для этого необходимо, чтобы после кроссинговера обе хромосомы (бывшие хроматиды каждого из гомологов) y+ sn отошли к одному полюсу клетки, а хромосомы y sn+ – к другому. Потомки дочерних клеток, размножившись на стадии куколки, и приведут к появлению мозаичных пятен. Таким образом, мозаичные пятна образуются тогда, когда рядом расположены две группы (точнее, два клона) клеток, фенотипически отличающиеся друг от друга и от клеток остальных тканей данной особи.
Неравный кроссинговер
Это явление было детально изучено на примере гена Bar (В – полосковидные глаза), локализованного в Х-хромосоме D. melanogaster. Неравный кроссинговер связан с дупликацией какого-либо участка в одном из гомологов и с утратой его в другом гомологе. Обнаружено, что ген В может присутствовать в виде тандемных, т. е. следующих друг за другом, повторов, состоящих из двух и даже трех копий. Цитологический анализ подтвердил предположение о том, что неравный кроссинговер может вести к тандемным дупликациям. В области, соответствующей локализации гена В, на препаратах политенных хромосом отмечено увеличение числа дисков, пропорциональное дозе гена. Предполагается, что в эволюции неравный кроссинговер стимулирует создание тандемных дупликаций различных последовательностей и использование их в качестве сырого генетического материала для формирования новых генов и новых регуляционных систем.
Регуляция кроссинговера
Кроссинговер – это сложный физиолого-биохимический процесс, который находится под генетическим контролем клетки и подвержен влиянию факторов внешней среды. Поэтому в реальном эксперименте о частоте кроссинговера можно говорить, имея в виду все те условия, в которых она была определена. Кроссинговер практически отсутствует между гетероморфными Х- и Y-хромосомами. Если бы он происходил, то хромосомный механизм определения пола постоянно разрушался бы. Блокирование кроссинговера между этими хромосомами связано не только с различием в их величине (оно наблюдается не всегда), но и обусловлено Y-специфичными нуклеотидными последовательностями. Обязательное условие синапса хромосом (или их участков) – гомология нуклеотидных последовательностей.
Для абсолютного большинства высших эукариот характерна примерно одинаковая частота кроссинговера как у гомогаметного, так и гетерогаметного полов. Однако есть виды, у которых Кроссинговер отсутствует у особей гетерогаметного пола, в то время как у особей гомогаметного пола он протекает нормально. Такая ситуация наблюдается у гетерогаметных самцов дрозофилы и самок шелкопряда. Существенно, что частота митотического кроссинговера у этих видов у самцов и самок практически одинакова, что указывает на различные элементы контроля отдельных этапов генетической рекомбинации в половых и соматических клетках. В гетерохроматических районах, в частности прицентромерных, частота кроссинговера снижена, и поэтому истинное расстояние между генами в этих участках может быть изменено.
Обнаружены гены, выполняющие функции запирателей кроссинговера, но есть также гены, повышающие его частоту. Они иногда могут индуцировать заметное число кроссоверов у самцов дрозофилы. В качестве запирателей кроссинговера могут выступать также хромосомные перестройки, в частности инверсии. Они нарушают нормальную конъюгацию хромосом в зиготене.
Обнаружено, что на частоту кроссинговера влияют возраст организма, а также экзогенные факторы: температура, радиация, концентрация солей, химические мутагены, лекарства, гормоны. При большинстве указанных воздействий частота кроссинговера повышается.
В целом кроссинговер представляет собой один из регулярных генетических процессов, контролируемых многими генами как непосредственно, так и через физиологическое состояние мейотических или митотических клеток. Частота различных типов рекомбинаций (мейотический, митотический кроссинговер и сестринские хроматидные обмены) может служить мерой действия мутагенов, канцерогенов, антибиотиков и др.
Биологическое значение кроссинговера
Благодаря сцепленному наследованию удачные сочетания аллелей оказываются относительно устойчивыми. В результате образуются группы генов, каждая из которых функционирует как единый суперген, контролирующий несколько признаков. В то же время, в ходе кроссинговера возникают рекомбинации – т.е. новые комбинации аллелей. Таким образом, кроссинговер повышает комбинативную изменчивость организмов.
Это означает, что…
а) в ходе естественного отбора в одних хромосомах происходит накопление «полезных» аллелей (и носители таких хромосом получают преимущество в борьбе за существование), а в других хромосомах скапливаются нежелательные аллели (и носители таких хромосом выбывают из игры – элиминируются из популяций)
б) в ходе искусственного отбора в одних хромосомах накапливаются аллели хозяйственно-ценных признаков (и носители таких хромосом сохраняются селекционером), а в других хромосомах скапливаются нежелательные аллели (и носители таких хромосом выбраковываются).
Эволюционное значение сцепленного наследования
В результате сцепления одна хромосома может содержать как благоприятные аллели (например, А), так и нейтральные или относительно неблагоприятные (например, N). Если некоторый гаплотип (например, AN) повышает приспособленность его носителей за счет наличия благоприятных аллелей A, то в популяции будут накапливаться как благоприятные аллели, так и сцепленные с ними нейтральные или относительно неблагоприятные N.
Пример. Гаплотип AN обладает преимуществом перед гаплотипом «дикого типа» (++) за счет наличия благоприятного аллеля А, и тогда аллель N будет накапливаться в популяции, если он селективно нейтральный или даже относительно неблагоприятный (но его отрицательное влияние на приспособленность компенсируется положительным влиянием аллеля А).
Эволюционное значение кроссинговера
В результате кроссинговера неблагоприятные аллели, первоначально сцепленные с благоприятными, могут переходить в другую хромосому. Тогда возникают новые гаплотипы, не содержащие неблагоприятных аллелей, и эти неблагоприятные аллели элиминируются из популяции.
Пример. Гаплотип Al оказывается неблагоприятным по сравнению с гаплотипом «дикого типа» (++) за счет наличия летального аллеля l. Поэтому аллель А (благоприятный, нейтральный ил несколько снижающий приспособленность) не может проявиться в фенотипе, поскольку данный гаплотип (Al) содержит летальный аллель l. В результате кроссинговера возникают рекомбинантные гаплотипы A+ и +l. Гаплотип +l элиминируется из популяции, а гаплотип A+ фиксируется (даже в том случае, если аллель А несколько снижает приспособленность его носителей).