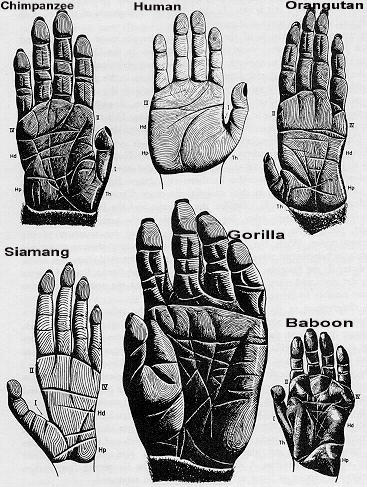
Фактори і критерії гомінізації
Рамки родини гомінід залежать від того, які ознаки лежать в основі визначення місця конкретних видів у системі гомінідів. Оскільки єдиним сучасним представником родини є людина, з її особливостей історично були виділені 3 найважливіші системи, що вважаються істинно гомінідними. Ці системи були названі гомінідною тріадою:
- прямоходіння (біпедія);
- кисть, пристосована до виготовлення знарядь;
- високорозвинений мозок.
До гомінідної тріади можна додати таку надійну ознаку, як маленькі ікла, які не виступають за лінію інших зубів.
Сучасні і викопні людиноподібні мавпи мають великі ікла, які виступають за лінію інших зубів. Серед інших типових гомінідних ознак ця з'явилася в еволюції раніше інших - початкова редукція іклів спостерігається вже у дріопітекових і сівапітекових приматів. Найдавніші гомініди і австралопітеки мали невеликі ікла, хоча розміри ікл австралопітеків виявляються майже посередині між розмірами у мавп і людини. Наскільки відомо на сьогоднішній день, всі чотириногі гоміноіди - володарі великих іклів, а прямоходячі - маленьких. Причини зменшення іклів залишаються неясними, але, ймовірно, криються в змінах типу харчування. Великі ікла, звичайно, потрібні приматам не для полювання, а для демонстрації сили хижакам і агресії родичам. Крім чисто захисної функції, ікла мають найважливіше комунікаційне призначення: за допомогою оскалу регулюються відносини, наводиться порядок у групах мавп, причому до контактної агресії справа майже ніколи не доходить, більше того, ця функція універсальна для більшості ссавців. В оригіналі це виглядає так: найбільший альфа-самець павіана сидить на найвищому і помітному місці і періодично скалить ікла. Інші мавпи бачать це і не скандалять між собою.
Зі зменшенням розмірів іклів в групах мавп повинна була виникнути проблема нерозуміння, оскільки демонстрація маленьких іклів далеко не настільки вражаюча. Мабуть, у цій недостатності експресії криються витоки підвищення інших комунікаційних способів, особливо міміки, жестикуляції і зародження мови - всі вони набагато краще розвинені у людини, ніж у приматів (звичайно, не можна абсолютизувати значення тільки цієї причини, були і багато інших). Таким чином, зменшення розмірів іклів мало грандіозні наслідки, і його не можна недооцінювати.
Прямоходіння визначається таким комплексом ознак:
Положення великого потиличного отвору - у прямоходячих знаходиться в центрі довжини основи черепа, відкривається вниз; у чотириногих - в задній частині основи черепа, повернене назад. Відповідно, основу черепа вкорочена у двоногих і видовжена у чотириногих. Варіант проміжного типу відомий вже у Sahelanthropus tchadensis близько 6-7 млн. років тому, а типовий для прямоходіння - у Ardipithecus ramidus ramidus 3,9-4,4 млн. років тому.
Будова таза - у прямоходячих таз широкий і низький; у чотириногих таз вузький, високий і довгий. Проміжний варіант виявляється у Ardipithecus ramidus 4,4 млн. років тому. Біпедальний варіант відомий, починаючи з Australopithecus afarensis 3,2 млн. років тому.
Будова довгих кісток ніг - у прямоходячих ноги довгі, тазостегнові суглоби сильно розведені один від одного внаслідок великої ширини тазу, а коліна зведені разом, так що стегнові кістки при погляді спереду нахилені, а кістки гомілки вертикальні, стопи зближені, при погляді збоку коліна випрямлені; у чотириногих приматів руки довші ніг, коліна розведені в сторони "колесом" і завжди напівзігнуті, стопи рознесені один від одної, так що при двоногому ходінні мавпа рухається дуже незграбно, похитуючись, компенсуючи нестійкість сильними бічними коливаннями тулуба. Обом варіантам відповідають певні характерні форми колінних і гомілковостопних суглобів. Біпедальна будова стегнових кісток відома, починаючи з Orrorin tugenensis 6,2 млн. років тому.
Будова стопи - у прямоходячих виражені поздовжній і поперечний склепіння (підйоми) стопи, пальці прямі, короткі, великий палець не відведений в бік і малорухомий; у чотириногих стопа пласка, пальці довгі, вигнуті, рухливі, великий палець має хапальну функцію, може сильно відводитися в сторону, що має відображення в більшій довжині його м'язів і характерній формі суглобів. У Ardipithecus ramidus 4,4 млн. років тому присутні склепіння стопи, але пальці довгі і зігнуті, а великий палець здатний відводитися далеко в бік. У стопі Australopithecus anamensis 4,1 млн. років тому, судячи з будови великої гомілкової кістки, великий палець був малорухомий. У Australopithecus afarensis 2,5-3,9 млн. років тому cводи стопи добре виражені, великий палець міг злегка протиставлятися іншим, але набагато слабкіше, ніж у сучасних мавп, відбиток ноги був майже як у сучасної людини. У стопі Australopithecus africanus і Paranthropus robustus великий палець був сильно відведений від інших, пальці були дуже рухливі, будова проміжна між мавпами і людиною. У всіх австралопітеків пальці стопи були досить довгі і зігнуті. У Homo habilis стопа сплощена, без вираженого зводу, але пальці прямі, короткі, а великий палець повністю приведений до решти.
Будова рук - у повністю прямоходячих гомінідів руки не пристосовані до ходіння по землі або лазання по деревах, руки короткі, фаланги пальців прямі; у різних приматів є численні морфологічні адаптації до чіпляння за гілки (включаючи варіанти з редукцією великого або інших пальців, або зі зрощенням пальців в єдиний "гачок"), а у вищих людиноподібних - до ходіння по землі з опорою на фаланги зігнутих пальців (включаючи особливу скошеність суглобової поверхні променевої кістки). Риси пристосування до ходіння по землі або лазіння по деревах є у австралопітеків Orrorin tugenensis, Ardipithecus kadabba, Ardipithecus ramidus, Australopithecus anamensis, Australopithecus afarensis, Australopithecus africanus, Paranthropus robustus і навіть Homo habilis. Зокрема, не виключено, що Australopithecus anamensis часто пересувалися на чотирьох з опорою на фаланги зігнутих пальців.
Будова хребта - у прямоходячих хребет орієнтований вертикально і має характерні вигини - лордози вперед і кіфози назад, розміри хребців закономірно збільшуються зверху-вниз, крижі широкі і короткі; у чотириногих немає шийного та поперекового лордозів, а розміри хребців розрізняються не настільки закономірно, крижі вузькі і довгі. У австралопітеків Australopithecus afarensis і Australopithecus africanus вигини, ймовірно, були як у сучасної людини, але деякі деталі будови хребців (наприклад, витягнутість тіла хребців спереду-назад) зближують їх з мавпами. Будова крижів у відомих австралопітеків - починаючи з Ardipithecus ramidus і Australopithecus afarensis - типово гомінідні.
Теорії походження прямоходіння
Міоценове похолодання
У середині і наприкінці міоцену в результаті глобального похолодання клімату відбулося значне скорочення площ тропічних лісів і збільшення площі саван. У таких умовах у численних жили тоді в тропічних лісах людиноподібних мавп намітилися три варіанти подальшої долі. Перший варіант - найсумніший, але найпоширеніший - вимирання: у цей час вимерла переважна більшість гоміноїдів. Другий - збереження в більш-менш незмінному вигляді у решти лісах: нащадками цих нечисленних приматів стали сучасні гібони, орангутанги, горили і шимпанзе. Третій варіант - перехід частини гоміноїдів до наземного способу життя у саванах, що розширювалися.
Трудова концепція
Згідно з широковідомою трудовою концепцією Ф. Енгельса і її пізнішими варіантами, виникнення прямоходіння тісно пов'язане зі спеціалізацією руки мавпи для трудової діяльності - перенесення предметів, дитинчат, маніпулювання їжею і виготовлення знарядь. У вульгарному викладі прямоходіння виникло для того, щоб звільнити руки для трудової діяльності. Надалі праця призвела до виникнення мови і суспільства. Однак, за сучасними даними, прямоходіння виникло набагато раніше виготовлення знарядь. Прямоходіння виникло не менше 6-7 мільйонів років тому у Sahelanthropus tchadensis і Orrorin tugenensis, а найдавніші знаряддя з Гони в Ефіопії мають датування лише 2,7 млн. років тому.
Втім, як "реінкарнацію" трудової теорії можна розглідати гіпотезу О.Лавджоя, згідно з якою прямоходіння з'явилося у зв'язку з особливою стратегією розмноження, через подовження дитинства і ослаблення межсамцевой агресії. Гомініди протягом дуже тривалого часу вирощують одного, максимум двох дитинчат. Безпомічних дитинчат треба носити, але в савані вигідніше не мати довгої шерсті, а охолоджуватися за допомогою потових залоз. Діти австралопітеків втратили можливість чіплятися за зниклу шерсть матерів, їх треба було притримувати руками. Разом з цим, батьки повинні були забезпечувати вимушено малорухомих самок їжею, а приносити її знову ж було зручніше в руках. А вже непрямим наслідком такої турботи набагато пізніше стало і використання знарядь праці. Згідно О.Лавджою, прямоходіння виникло ще в тропічному лісі, а в савани переселилися вже двоногі гомініди.
Інші теорії походження прямоходіння
Значна більшість інших гіпотез пов’язують прямоходіння зі зміною життя на деревах на наземне життя. Біпедія могла виникнути для орієнтування в савані, коли треба було дивитися поверх високої трави для пошуку їжі або виявлення хижаків. Крім того, прямоходячий індивід має психологічну перевагу перед чотириногим тваринам, оскільки дивиться зверхньо і так здається більшим і сильнішим, хоча може і не мати реального підтвердження своєї могутності. Піднімання рівня погляду для посилення враження на супротивника є універсальним способом серед усіх хребетних тварин, оскільки в світі тварин працює принцип "хто вищий, той головніший". Для приматів у савані цей аспект прямоходіння міг мати важливе значення.
Крім того, експериментально і на математичних моделях доведено, що двоноге пересування на великі відстані з середньою швидкістю енергетично більш вигідне, ніж чотириноге. У савані двоногий примат нагрівається значно менше чотириногого, оскільки під сонячні промені потрапляють тільки голова і плечі, а не вся спина, що особливо актуально поблизу екватора.
Згідно так званої "гіпотези водної мавпи", дуже детально розробленої Я.Ліндбландом, предки людини могли вставати на задні ноги, щоб переправлятися через водні перешкоди. Відомо, що більшість австралопітеків мешкали недалеко від води і, ймовірно, добували у воді частину свого прожитку. У будові людини є ряд ознак, що свідчать про значну адаптацію людини до плавання і пірнання, на відміну від орангутанів, горил і шимпанзе: положення волосся на тілі у напрямку від маківки до ніг - за течією води при пірнанні, орієнтація ніздрів вниз - для збереження повітря в носовій порожнині, здатність затримувати подих, редукований волосяний покрив на тілі, неекономне витрачання води організмом, що вкрай нетипово для тварин савани, невеликі перетинки між пальцями, небоязнь води. "Гіпотеза водної мавпи" в своєму крайньому варіанті багаторазово піддавалася критиці, але деякі її положення не можна ігнорувати.
Отже, прямоходіння виникло не з однієї якоїсь причини, а під впливом цілого комплексу передумов.
Кисть, пристосована до виготовлення і використання знарядь
Тут можна виділити такий трудовий комплекс:
Сильне зап'ястя. У австралопітеків, починаючи з Ardipithecus ramidus і Australopithecus anamensis, будова зап'ястя проміжна між мавпами і людиною. Майже сучасна будова спостерігається у Homo habilis 1,8 млн. років тому.
Протиставлення великого пальця кисті. Ознака відома вже 4,4 мільйона років тому у Ardipithecus ramidus і 3,2 мільйона років тому у Australopithecus afarensis і Australopithecus africanus. Протиставлення було повністю розвинене у Paranthropus robustus і Homo habilis 1,8 мільйона років тому. Нарешті, було своєрідним або обмеженим у неандертальців Європи близько 40-100 тисяч років тому.
Широкі, сплощені і вкорочені кінцеві фаланги пальців. Дуже широкі фаланги були у Paranthropus robustus, Homo habilis і всіх більш пізніх гомінідів. Цікаво, що Paranthropus robustus за цією ознакою навіть еволюційно обганяв "людину умілу" Homo habilis, хоча нашим предком була саме остання.
Прикріплення м'язів, що рухають пальці, майже сучасного типу відмічено у Paranthropus robustus і Homo habilis, але у них є і примітивні риси.
Кістки кисті у найдавніших прямоходячих гоміноїдів (Orrorin tugenensis, Ardipithecus kadabba, Ardipithecus ramidus, Australopithecus anamensis і Australopithecus afarensis) мають суміш ознак людиноподібних мавп і людини. Мабуть, ці види могли використовувати предмети як знаряддя, але не виготовляти їх. Перші виробники справжніх знарядь - Homo habilis. Можливо, знаряддя виготовляли і південноафриканські масивні австралопітеки Paranthropus robustus.
Трудова кисть в цілому сформувалася близько 1,8 мільйонів років тому.
Високорозвинений мозок
Мозок сучасної людини сильно відрізняється від мозку людиноподібних мавп за розмірами, формою, будовою і функціями, однак серед викопних форм можна знайти безліч перехідних варіантів. Типові ознаки мозку людини такі:
Великі загальні розміри мозку. У австралопітеків розмір мозку був як у сучасних шимпанзе - близько 320-550 см3. Бурхливе збільшення розмірів відбулося у Homo habilis близько 2,5-1,8 млн. років тому, а у пізніших гомінідів спостерігається плавне збільшення до сучасних значень.
Специфічні поля мозку. Зони Брока, Верніке (мовленнєві центри) та інші почали розвиватися у Homo habilis і архантропів, але повністю сучасного вигляду досягли, мабуть, тільки у сучасної людини.
Будова часток мозку. У людини значно розвинені лобні частки і нижня тім'яна часточка, скронева і лобова частки сходяться під гострим кутом, скронева частка широка й округла спереду, потилична частка відносно невелика, нависає над мозочком. У австралопітеків будова і розміри мозку були такі ж, як у людиноподібних мавп. Будова мозку Homo habilis була примітивною, але сам мозок мав порівняно великі розміри. Для пізніших гомінідів характерне плавне наближення до сучасної будови мозку.
Гомінідний тип мозку виник близько 1,8 млн. років тому або дещо раніше, причому збільшенню розмірів передувало його ускладнення. Надалі мозок активно еволюціонував, а повністю сучасний мозок сформувався лише близько 25 тисяч років тому.
Антропогенез, будучи, з одного боку, цілком закономірним і навіть передбачуваним епізодом у розвитку органічної матерії, епізодом зумовленим її базовими властивостями і самими умовами земного існування, виявився в той же час, як це не парадоксально, результатом цілого ряду збігів, наслідком далеко не обов'язкового і навіть малоймовірного перетину в одному місці і в один час незалежних чи дуже мало залежних один від одного природних процесів. Це в рівній мірі відноситься і до пізніх етапів антропогенезу, коли вирішувалося питання про те, якому з видів гомінід володіти в майбутньому Землею, і до самого початкового його періоду, коли гілку, що вела до людини, було ще практично неможливо розрізнити на генеалогічному дереві приматів, і наші предки робили лише перші кроки в напрямку гомінізаціі. Поява людини взагалі і виду Homo sapiens, зокрема, може, таким чином, розглядатися, з одного боку, як найбільш яскравий прояв «магістральної» тенденції в розвитку живої природи на Землі, а з іншого, як випадковість, що відбулася завдяки унікальному збігові настільки ж унікальних обставин.
http://beday.ru/9.shtml
http://antropogenez.ru/zveno/p/2/