

2. Типи визначення статі.
В залежності від того, в якій період онтогенезу визначається стать, розрізняють наступні типи встановлення статі:
1. Прогамний тип (до запліднення) характерний для видів, у яких гетерогаметною є жіноча стать. У них вже в незаплідненій яйцеклітині закладена генетична інформація щодо статі майбутнього нащадка.
2. Сингамний тип характерний для видів, у яких жіноча стать гомогаметна і стать нащадків визначається хромосомним набором чоловічої гамети.
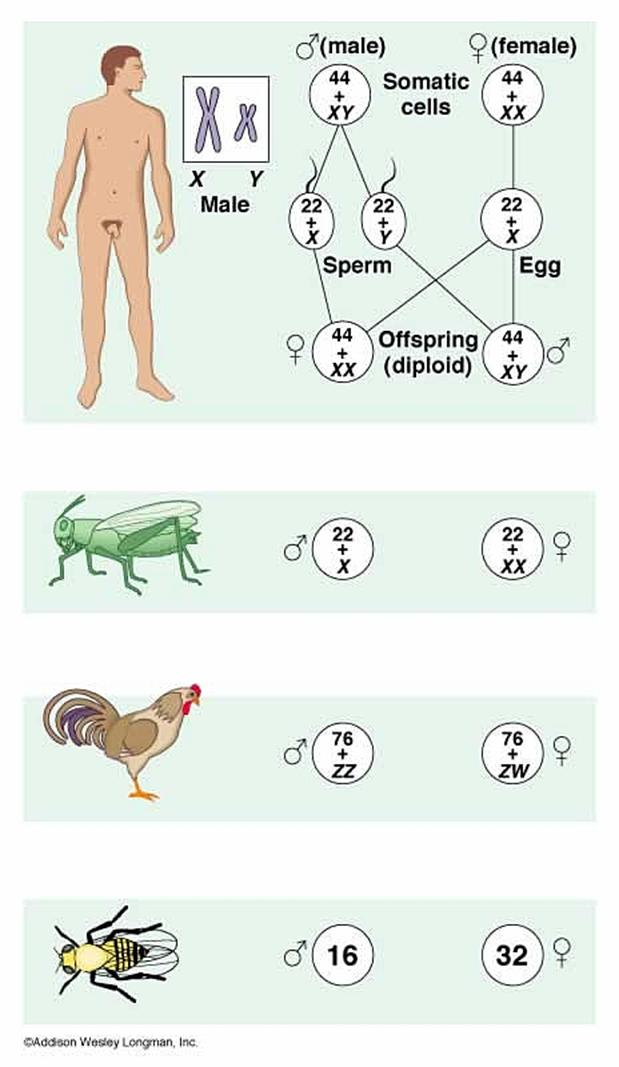
Рисунок 3. Хромосомний механізм визначення статі
3. Епігамний тип визначення статі характеризується тим, що формування статевих ознак в онтогенезі відбувається під впливом зовнішніх факторів.
У морського черва Bonellia viridis відсутні статеві хромосоми. Так, рухомі личинки Bonellia viridis розвиваються в самок, якщо прикріплюються до субстрату (рис. 4а). Якщо ж личинка прикріплюється до самки, вона розвивається у рудиментарного самця, який веде напівпаразитичний спосіб життя.


а б
Рис. 4а. Морський черв Bonellia viridis, 4б – черепаха
У деяких рептилій, наприклад, крокодилів, деяких ящірок і черепах (рис. 4б) майбутня стать визначається температурою, при якій розвиваються ембріони в яйцях.
Температурна залежність може визначатися різними кривими. Чоловіча стать може розвиватися за низьких або високих температур, або за оптимальних. а відхилення температурв одну чи іншу сторону призводить до розвитку жіночої статі (рис. 5). Температурні межі, як правило, незначні, а саме відхилення на 1-5 оС впливає на визначення статі.
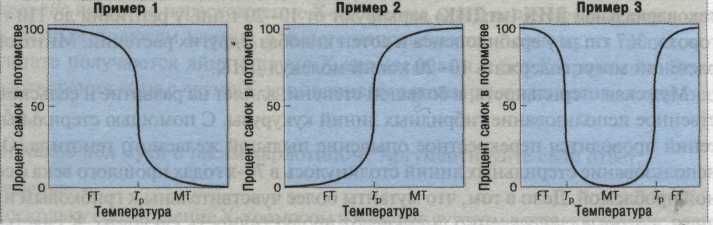
Рис. 5. Три різних спектра температурозалежної детермінації статі у рептилій
3. Балансова теорія визначення статі.
Простий з першого погляду хромосомний механізм визначення статі у дрозофіли виявився значно складнішим. Вивчення мух із аномальним набором статевих хромосом показано, що Y – хромосома не має істотного впливу на розвиток статі. Так, особини із каріотипом 2А (набор аутосом) + ХО є типовими, але безплідними, самцями, а особини із каріотипом 2А+ХХY, незважаючи на наявність Y – хромосоми, фертильними самками (табл.1).
К.Бріджес висловив припущення, що у дрозофіли гени чоловічих потенцій знаходяться в аутосомах, а жіночих – у Х – хромосомі, і що визначення статі залежить від співвідношення кількості аутосом і Х – хромосом. У дослідах на дрозофілі його гіпотеза про те, що стать визначається балансом генів аутосом і Х – хромосом, підтвердилась. Однак у більшості тварин ця закономірність не простежується.
У людини, наприклад, чоловічу стать незалежно від кількості Х – хромосом визначає наявність Y – хромосоми. Людина із набором статевих хромосом ХХУ буде фенотипово чоловіком, хоча із певними порушеннями (синдром Кляйнтфельтера), а із набором статевих хромосом Х0 буде фенотипово жінкою, хоча стерильною із розумовим відставанням (синдром Шерешевського-Тернера).
Таблиця 1.
Вплив співвідношення аутосом і Х – хромосом на вияв статі у дрозофіли
3Х+2А |
3:2 |
Надсамка |
3Х+3А |
1:1 |
Самка |
2Х+2А |
1:1 |
Самка |
У+2Х+2А |
1:1 |
Самка |
2Х+3А |
2:3 |
Інтерсекс |
2Х+У+3А |
2:3 |
Інтерсекс |
Х+2А |
1:2 |
Самець |
Х+У+2А |
1:2 |
Самець |
Х+3А |
1:3 |
Надсамець |
Хромосомний механізм визначення статі у рослин поділяється на два основних типи:
У одних рослин у визначенні чоловічої статі активну роль виконує Y – хромосома і навіть у тетраплоїда ХХХY – стать чоловіча. Лише наявність чотирьох Х – хромосом (ХХХXY) зумовлює розвиток гермафродитних рослин. Прикладом таких рослин слугує Melandrium alba (куколиця біла), Brionia dioica (переступень дводомний).
У інших рослин стать визначається балансом аутосом і Х – хромосом, при цьому Y – хромосома практично інертна. Прикладом таких рослин слугує Rumex acetosa (щавель кислийта).
Отже,стать не завжди визначається співвідношенням генів аутосом і гетерохромосом, тому балансова теорія К.Бріджеса вимагає деяких уточнень.