

Хугстеновское спаривание – основа формирования триплексов днк. Н-днк.
В крупных хромосомах обнаружен ряд структурных вариантов двойной спирали, отличных от описанных выше, которые определяются специфическими последовательностями нуклеотидов в ДНК. Эти специфические последовательности способны влиять на функцию и разнообразные ДНК-зависимые процессы в тех участках ДНК, которые расположены в непосредственной близости от таких последовательностей. Например, в двойной спирали ДНК формируются своеобразные изгибы, если в одной цепи присутствуют четыре или более адениновых нуклеотидов. Шесть адениновых нуклеотидов расположенных подряд в одной цепи приводят к отклонению центральной оси в ДНК на 18. Наблюдаемые изгибы двойной спирали, индуцируемые определенными последовательностями нуклеотидов, могут играть существенную роль в связывании ряда белков с ДНК.
Наиболее известными и часто встречающимися в ДНК последовательностями являются, так называемые, палиндромы. Палиндромом называют слово, фразу или даже изречение, которое читается слева направо и наоборот одинаково. Примерами таких слов или фраз являются: ШАЛАШ, КАЗАК, ПОТОП (ROTATOR), А РОЗА УПАЛА НА ЛАПУ АЗОРА (NURSES RUN). Данный термин применяют к тем регионам ДНК, которые характеризуются наличием инвертированных повторов последовательностей оснований имеющих симметрию второго порядка относительно двух цепей ДНК (рис. 3.9). Такие последовательности, по вполне понятной причине, являются самокомплементарными и имеют склонность к образованию шпилечных или крестообразных структур (рис. 3.10).
В тех случаях, когда инвертированный повтор присутствует в одной и той же цепи ДНК такая последовательность называется зеркальным повтором. Зеркальные повторы не обладают свойствами самокомплементарности и, поэтому не способны к формированию шпилечных или крестообразных структур. Последовательности такого типа обнаружены практически во всех крупных молекулах ДНК и могут включать от всего нескольких пар оснований до нескольких тысяч пар оснований.
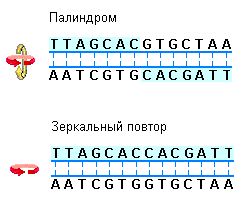
Рис. 3.9 |
Палиндромы и зеркальные последовательности. Палиндромы представляют собой специфические последовательности в двухцепочечной ДНК, которые обладают симметрией второго порядка. Для того, чтобы наложить один повтор на другой его необходимо сначала повернуть на 180 относительно горизонтальной оси, а затем на 180 относительно вертикальной оси (как это показано цветными стрелками). Иными словами симметрия второго порядка означает, что при повороте двухцепочечной последовательности на 180 относительно оси перпендикулярной к плоскости, в которой расположены основания, их положения в двух цепях ДНК не изменятся. С другой стороны, зеркальный повтор отличается симметрией в каждой цепи ДНК. Для наложения одного повтора на другой достаточно одного единственного поворота последовательности на 180 вокруг вертикальной оси (Nelson D.L., Cox M.M., Lehninger Principles of Biochemistry, W.H. Freeman (ed.), San Francisco, 2004). |
Присутствие палиндромов в виде крестообразных структур в эукариотических клетках не доказано, хотя некоторое количество крестообразных структур обнаружено в условиях in vivo в клетках E. coli. Наличие в составе РНК или одноцепочечных ДНК самокомплементарных последовательностей служит основной причиной фолдинга в растворах сложных структур отличающихся формированием множества «шпилек».

Рис. 3.10 |
Шпильки и крестообразные структуры. Палиндромные последовательности в ДНК (или РНК) способны формировать альтернативные структуры за счет внутрицепочечного спаривания оснований. (а) – Шпилечные структуры характерны, в первую очередь, для одноцепочечных нуклеиновых кислот, т.е. для РНК. (b) – Участки двойной спирали ДНК содержащие палиндромы способны образовывать крестообразные структуры (Nelson D.L., Cox M.M., Lehninger Principles of Biochemistry, W.H. Freeman (ed.), San Francisco, 2004). |
В последнее время было описано несколько необычных вариаций строения ДНК, которые могут включать трех- и четырех цепочечные структуры. Кроме описанных выше форм ДНК и специфических последовательностей оснований, ответственных за образование, например, крестообразных структур к настоящему времени удалось охарактеризовать несколько совсем необычных структур, которые построены из трех или даже четырех цепей ДНК. Указанные структурные вариации заслуживают пристального внимания со стороны исследователей по той причине, что имеет место определенная тенденция участия подобных структур в инициации или регуляции ДНК-зависимых процессов (репликации, рекомбинации и транскрипции). Показано, что нуклеотиды, принимающие участие в классическом уотсон-криковском спаривании оснований способны образовывать ряд дополнительных водородных связей особенно с функциональными группами, расположенными в большой бороздке ДНК. Например, остаток протонированного цитидина может спариваться с остатком гуанозина в составе пары G ≡ C, а тимидин спаривается с аденозином в составе пары А = Т (рис. 3.11). Положения в пуринах атомов N-7, O6 и N6, которые принимают участие в образовании водородных связей в составе триплексов ДНК, часто называют положениями Хугстена, а не уотсон-криковское спаривание называют хугстеновским спариванием по имени Карста Хугстена впервые описавшего в 1963 году возможность подобного спаривания. Именно хугстеновское спаривание оснований лежит в основе формирования триплексов ДНК.

Рис. 3.11 |
Структуры, построенные из трех и четырех цепей ДНК. (а) – Способы спаривания оснований в наиболее хорошо охарактеризованных триплексах ДНК. Хугстеновское спаривание оснований в каждом случае показано красным цветом. (b) – Трехспиральная ДНК, состоящая из двух пиримидиновых цепей (поли-Т) и одной пуриновой цепи (поли-А) (PDB ID 1BCE). Темно-синие и светло-синие цепи антипараллельны и спарены по правилу Уотсона-Крика. Третья цепь (состоящая только из пиримидинов) параллельна пуриновой цепи и спарена за счет водородных связей хугстеновского взаимодействия. Тройная спираль показана с торца и включает пять триплексов. (с) – Способ спаривания гуанозинов в четырехспиральной ДНК. (d) – Показаны два тетраплета, состоящие их четырех остатков гуанозина (PDB ID 1QDG). (е) – Возможные варианты ориентации цепей в четырехспиральной ДНК, построенной из тетраплетов G (Nelson D.L., Cox M.M., Lehninger Principles of Biochemistry, W.H. Freeman (ed.), San Francisco, 2004). |
Триплексы, приведенные на рис. (а, b) отличаются наибольшей стабильностью при низких значениях рН поскольку образование, например, триплета C ≡ G ∙ С+ возможно в случае протонирования цитидина (см. рис. 3.11). В таком триплексе значение рКа для этого цитидина составляет > 7,5 в отличие от его нормального значения 4,2. Еще одна особенность триплексов заключается в том, что они достаточно легко образуются в тех случаях, когда в ДНК присутствуют протяженные участки, состоящие только из пиримидинов или пуринов в одной цепи. Некоторые триплексные ДНК могут состоять из двух пиримидиновых цепей и одной пуриновой или, наоборот, двух пуриновых цепей и одной пиримидиновой.
Возможно также спаривание четырех цепей ДНК с образованием тетраплексов, но этот процесс идет относительно легко, только если в ДНК имеются последовательности, содержащие очень большое количество остатков гуанозина. Гуанозиновые тетраплексы или иначе G-тетраплексы весьма стабильны в широком диапазоне физиологических условий в клетке.
Наиболее экзотической структурой ДНК является так называемая Н-ДНК, которая обнаруживается в полипиримидиновых или полипуриновых участках содержащих зеркальные повторы. Известным примером последовательности склонной к образованию такой структуры являются протяженные участки в ДНК содержащие попеременно следующие друг за другом остатки Т и С (рис. 3.12). Характерной структурной особенностью Н-ДНК является ее трехцепочечная форма, показанная на рис. 3.12 (а, b). Две из трех цепей в тройной спирали Н-ДНК содержат пиримидины и одна цепь состоит из пуринов.
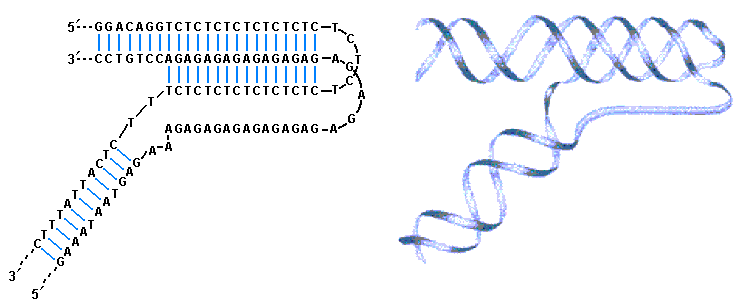
Рис. 3.12 |
Структура Н-ДНК. (а) – Последовательность в цепи ДНК, которая представляет собой попеременно расположенные остатки Т и С можно, в принципе, рассматривать как зеркальный повтор относительно остатка Т или остатка С. (b) – Такие последовательности способны участвовать в формировании необычной структуры, в которой две цепи одной части зеркального повтора в ДНК разделяются и цепь содержащая пиримидины другой части зеркального повтора разворачивается так, что это создает условия для образования триплекса. При этом цепь содержащая пурины (попеременно расположенные остатки А и G) в другой части повтора остается не спаренной. Эта структура образует острый изгиб в молекуле ДНК (Nelson D.L., Cox M.M., Lehninger Principles of Biochemistry, W.H. Freeman (ed.), San Francisco, 2004). |
В ДНК живых клеток сайты узнаваемые многими ДНК-связывающими белками организованы в виде палиндромов, полипиримидиновых или полипуриновых последовательностей, которые могут образовывать тройные спирали или даже Н-ДНК. Указанные структуры обнаруживаются в областях ДНК, ответственных за регуляцию экспрессии ряда генов у эукариот. Установлено, что синтетические полинуклеотиды способные формировать тройные спирали могут выступать в роли потенциальных ингибиторов экспрессии генов. В свою очередь подобная активность трехцепочечных ДНК может найти применение в медицине и сельском хозяйстве.