

Принципиальная организация днк. Модель днк Уотсона и Крика
Открытие двойной спирали ДНК было одним из самых выдающихся событий молекулярной биологии. В основу модели ДНК, построенной в 1953 году Джеймсом Уотсоном и Фрэнсисом Криком, легли две группы фактов: во-первых, результаты предшествующего анализа количественного соотношения четырех оснований в составе ДНК из разных источников, полученные Эрвином Чаргаффом и, во-вторых, анализ картин дифракции рентгеновских лучей на нитях ДНК, полученных Розалиндой Франклин и Морисом Уилкинсом.
Закономерности, выявленные Э.Чаргаффом были впоследствии сформулированы в виде правил [7]:
а) состав ДНК различных клеток, составляющих ткани и органы одного организма всегда одинаков;
б) состав ДНК клеток организма с возрастом не изменяется;
в) состав ДНК клеток разных видов различен;
г) количество аденина всегда равно количеству тимина (АТ), а количество гуанина равно количеству цитозина (G C);
д) сумма пуриновых оснований равна сумме пиримидиновых оснований (А+G Т+С).
При этом, любая молекула ДНК может быть охарактеризована по ее составу, как отношение (G+C)/(A+T), которое изменяясь от 26 до 74 мольных % остается характерным для каждого вида.
Данные рентгеноструктурного анализа показали, что ДНК имеет форму регулярной спирали диаметром 20 Å, делающей один полный оборот каждые 34 Å (3,4 нм). Поскольку расстояние между соседними нуклеотидами в цепи составляло 3,4 Å (0,34 нм), то в одном витке должно быть 10 нуклеотидов.
Плотность ДНК свидетельствовала о том, что спираль должна состоять из двух полинуклеотидных цепей. Одновременно, постоянный диаметр спирали предполагал, что в каждой цепи азотистые основания направлены внутрь спирали, причем расположены так, что пуриновое основание взаимодействует с пиримидиновым, обеспечивая ситуацию, при которой невозможно образование контактов пурин-пурин и пиримидин-пиримидин.
Дж. Уотсон и Ф. Крик предположили, что две полинуклеотидные цепи в молекуле ДНК не связаны ковалентно, а соединяются водородными связями, возникающими между азотистыми основаниями [8]. Реакции взаимодействия G с C и A с T получили название спаривания оснований, а основания, способные образовывать пары, получили название комплементарных. В А-Т паре основания соединены двумя водородными связями: одна из них образуется между амино- и кето-группами, а другая – между двумя атомами азота пурина и пиримидина, соответственно. В G-C паре имеются три водородные связи: две из них образуются между амино- и кето-группами соответствующих оснований, а третья – между атомами азота (рис. 3.3).
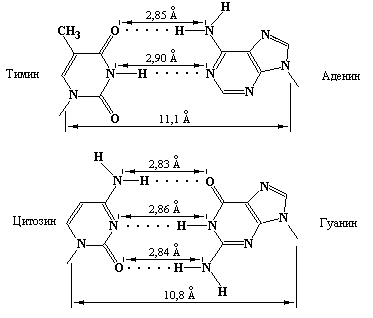
Рис. 3.3 Водородные связи между комплементарными основаниями. Как следует из рисунка, геометрия двух типов пар оснований практически одинакова (Kornberg A, DNA replication, Freeman W.H. (ed.), San Francisco, 1980).
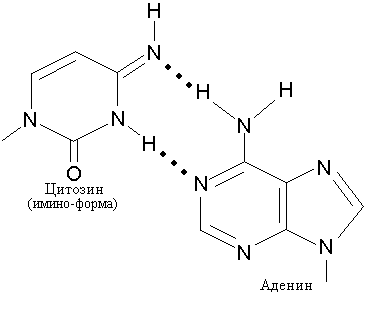
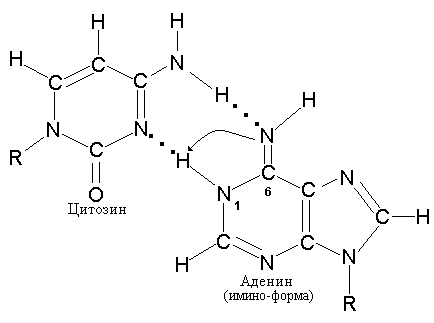
Для осуществления специфического спаривания основания должны находиться в соответствующей таутомерной форме. Миграция водородного атома позволяет каждому основанию существовать в различных таутомерных формах. Основания, входящие в состав двойной спирали ДНК и образующие канонические пары, должны иметь амино-группы (-NH2) и кето-группы (СО) в отличие от таутомеров, имеющих имино-группы (NH) и енольные группы (-ОН) и способных к неканоническому спариванию, как, например, пуриновое основание аденин, который может образовать пару с таутомерной формой цитозина рис. 3.4).
А В
Рис. 3.4 Образование неканонических пар с участием имино-формы цитозина и аденина (А) и имино-формы аденина с нормальным цитозином (В).
Действительно, образование пар между двумя пуринами, двумя пиримидинами или некомплементарными основаниями A-C или G-T стерически затруднено, поскольку при этом не могут образовываться подходящие водородные связи и, следовательно, нарушается геометрия спирали. Модифицированные пурины и пиримидины, с небольшой частотой встречающиеся в ДНК, образуют такие же водородные связи, что и их немодифицированные аналоги. В этом случае правило спаривания не нарушается.
Согласно модели двойной спирали две полинуклеотидные цепи в молекуле ДНК - антипараллельны, т. е. идут в противоположных направлениях. Поэтому, рассматривая спираль вдоль оси, можно видеть, что одна цепь идет в направлении 5/ 3/, а другая - в направлении 3/ 5/ (рис. 3.5).
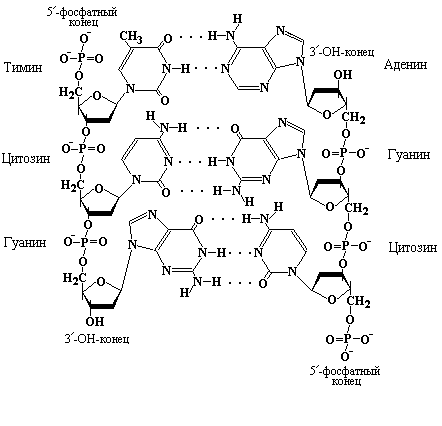
Рис. 3.5 Двойная спираль ДНК имеет постоянный диаметр, поскольку пурины расположены напротив пиримидинов, образуя канонические пары A-T и G-C. Сахарофосфатный остов спирали расположен снаружи. Одна полинуклеотидная цепь имеет направление 5/ 3/, а другая – 3/ 5/. Рисунок схематичен и закручивание цепей в двойную спираль не показано.
Основания имеют плоскую форму и располагаются парами в плоскости, перпендикулярной оси спирали. Вдоль спирали основания уложены стопками друг на друга и стабилизация спиральной структуры дополнительно обеспечивается межплоскостными взаимодействиями между ароматическими кольцами соседних оснований. Эти специфические контакты получили название стэкинг-взаимодействий, которые являются результатом реализации вандерваальсовых сил, возникающих за счет перекрывания -облаков над и под двойными связями ненасыщенных колец пуринов и пиримидинов, с одной стороны, и гидрофобных взаимодействий, с другой стороны.
Каждая пара оснований в ДНК повернута на 36 вокруг оси спирали относительно следующей пары и, таким образом, 10 пар оснований составляют один полный оборот в 360. Две цепи, образующие двойную спираль, уложены таким способом, что наблюдаемая структура характеризуется наличием малой бороздки шириной 12 Å и большой бороздки шириной 22 Å. Двойная спираль правосторонняя: если смотреть вдоль оси спирали – повороты следуют по часовой стрелке. Данное описание соответствует модели ДНК, известной как B-форма (рис. 3.6).
Одно из обязательных требований к генетическому материалу состоит в том, что он должен быть способен к точному самовоспроизведению при каждом клеточном делении. По этому поводу в своих пионерских работах Дж. Уотсон и Ф. Крик сделали классический вывод: «Нельзя не заметить, что постулированное нами специфическое взаимодействие непосредственно предполагает возможность копирования генетического материала».

Рис. 3.6 Пространственные модели В-, А- и Z-форм ДНК (каждая модель содержит 20 пар оснований). Атомы фосфора и связанные с ними атомы кислорода показаны в виде темных шариков. Сплошные линии, соединяющие фосфатные группы, показывают ход полинуклеотидных цепей. Показательна зигзагообразная форма остова Z-ДНК. А-ДНК короче и толще, а Z-ДНК несколько длиннее и тоньше, чем В-ДНК (Сингер М., Берг П., Гены и геномы, М., «Мир», 1998, т. 1, с. 44).
Эта идея базировалась на том представлении, что соединение двух полинуклеотидных цепей только водородными связями должно позволять им разделяться.
Предлагая модель молекулы ДНК Дж. Уотсон и Ф. Крик постулировали также механизм репликации ДНК, в соответствии с которым каждая исходная цепь может служить матрицей для синтеза комплементарной дочерней цепи. Такой механизм удвоения ДНК был назван полуконсервативным.
Вращение связей между атомами, формирующими сахарофосфатный остов ДНК и свободное вращение С-1/-N-гликозидной связи – основа структурных вариаций в ДНК. Cин- и анти-конформации.
Альтернативные двухспиральные структуры ДНК. Параметры B-, A- и Z-форм ДНК.
Существовавшее до недавнего времени мнение о том, что В-форма ДНК – это совершенная двойная спираль, геометрия которой одинакова независимо от нуклеотидной последовательности, в действительности не совсем корректно. Детальный рентгено-структурный анализ, построение моделей и термодинамические расчеты показали, что плоскости соседних пар оснований не строго параллельны. Каждая комплементарная пара оснований является как бы клином, отклоняющим ось спирали в одном или другом направлении. Наибольшее «отклонение» наблюдается тогда, когда два соседних аденина в одной цепи спарены с двумя тиминами другой. В этом месте происходит локальное искривление спирали. Если такие пары встречаются с периодичностью примерно один раз на 10 пар нуклеотидов (т.е. один раз на каждый виток спирали), то молекула ДНК приобретает заметно искривленную форму. Изгибы в молекуле ДНК наблюдаются в тех участках последовательности, где с необычно высокой частотой встречаются повторы [А-Т]5-6, разделенные GC-богатыми участками из четырех-шести нуклеотидов. Биологическая роль искривления двойной спирали ДНК окончательно не установлена. Предрасположенность к такому изгибанию, зависящая от последовательности оснований, может иметь значение при наматывании молекулы ДНК на гистоновые октамеры в хроматине. Возможно, изгибание ДНК существенно и при специфическом связывании ДНК с белками в процессе регуляции экспрессии генов.
Таким образом, стало очевидным, что некоторые параметры классической B-формы спирали необходимо пересмотреть. Анализ моделей позволил также сделать предположение о том, что ДНК может образовывать другие типы двуспиральных структур.
Проблема уточнения характеристик двойной спирали связана с тем, что классические параметры были получены методом дифракции рентгеновских лучей на волокнах ДНК. Однако этим методом нельзя определить положение отдельных атомов. До сих пор мы говорили о ДНК как о длинной жесткой двойной спирали, но в настоящее время известно, что она должна быть свернута и плотно уложена для того, чтобы поместиться в ядре клетки. Естественно, что при этом отдельные детали ее структуры могут изменяться.
По крайней мере сейчас не вызывает сомнения, что ДНК - это всегда двойная спираль.
В результате уточнений оказалось, что число пар оснований в одном витке не 10,0, а 10,5. Это различие вызвало необходимость изменить угол вращения между соседними парами оснований до 34,6, таким образом, отрезок спирали в пределах которого совершается полный оборот в 360 стал несколько длиннее. Особенно важно, что величина 10,5 пар оснований является средним значением для целой молекулы ДНК в определенных условиях.
Ввиду таких вариаций идея о существовании единственной структуры сменилась представлениями, допускающими наличие семейства двухспиральных структур.
Действительно, несмотря на кажущуюся жесткость структуры, ДНК является удивительно гибкой и подвижной молекулой. Благодаря заметному вращению связей участвующих в формировании сахаро-фосфатного остова ДНК, термическим колебаниям эта макромолекула склонна к изгибанию, растягиванию, а также плавлению (разделению комплементарных цепей). В настоящее время уже известно множество структурных вариантов ДНК обнаруживаемых в живых клетках, которые значительно отличаются от классической уотсон-криковской структуры, при этом некоторые из них или даже все структуры играют определенную роль во многих ДНК-зависимых процессах. Важно, что эти разнообразные структурные варианты абсолютно не противоречат основополагающим принципам строения ДНК, сформулированным Уотсоном и Криком: комплементарности цепей, их антипараллельности и правилам спаривания оснований – А = Т и G ≡ C.
Структурные вариации в ДНК могут зависеть от трех основных процессов: изменения конформации остатков дезоксирибозы, вращения связей между атомами, формирующими сахарофосфатный остов ДНК и свободного вращения С-1/-N-гликозидной связи (рис. 3.7а). Вследствие стерических причин пуриновые основания в составе пуриновых нуклеотидов в ДНК могут приобретать относительно остатка дезоксирибозы две стабильные конформации обозначаемые как син и анти (см. рис. 3.7b). В то же время пиримидиновые основания пиримидиновых нуклеотидов присутствуют в ДНК в виде анти-конформеров, что связано со стерическими несоответствиями, возникающими между углеводной частью нуклеотида и карбонильным кислородом в С-2 положении пиримидина (см. рис. 3.7b).


Рис. 3.7 |
Вариации структуры ДНК. (а) – Конформация нуклеотида в составе ДНК может изменяться за счет вращения любой из семи связей показанных на рисунке. Шесть связей вращается свободно. Ограниченное, но все же возможное вращение вокруг связи 4 приводит к такому искажению кольца, при котором один из атомов пятичленного фуранозного кольца оказывается вне плоскости создаваемой четырьмя другими атомами. Такая конформация представляет собой эндо- или экзо-структуру, в зависимости от того располагается ли данный атом на той же стороне плоскости, что и С-5/ или на противоположной стороне (рис. 3.8). (b) – Пуриновые основания в составе соответствующих нуклеотидов могут принимать только две стерически доступные конформации – син или анти относительно характера присоединения основания к остатку дезоксирибозы. Пиримидиновые основания, главным образом, приобретают анти-конформацию (Nelson D.L., Cox M.M., Lehninger Principles of Biochemistry, W.H. Freeman (ed.), San Francisco, 2004). |
Нужно еще раз подчеркнуть, что классическая модель ДНК Уотсона-Крика, о которой шла речь выше, и которая получила название В-формы ДНК или, просто, В-ДНК является наиболее стабильной структурой хорошо описывающей в физиологических условиях строение этой двухцепочечной молекулы, не имеющей специфических последовательностей нуклеотидов. Именно по этой причине В-форма ДНК является структурой сравнения для изучения и описания многих других форм молекул ДНК. В настоящее время известны два хорошо описанных варианта ДНК, полученных с помощью рентгеноструктурного анализа – это А- и Z-формы ДНК. Характерные особенности строения В-, А- и Z-форм ДНК и их отличия суммированы в таблице 1.
Таблица 1
Параметры спирали
|
А-форма |
В-форма |
Z-форма |
Направление спирали |
Правосторонняя |
Правосторонняя |
Левосторонняя |
Диаметр |
26 Ǻ |
20 Ǻ |
18 Ǻ |
Число пар оснований в одном витке спирали |
11 |
10,5 |
12 |
Расстояние между па-рами оснований по ходу спирали |
2,6 Ǻ |
3.4 Ǻ |
7,7 Ǻ |
Наклон плоскости в которой расположены основания по отношению к перпендикуляру проведенному к оси спирали |
20 |
6 |
7 |
Конформация углевода |
С-3/ эндо |
С-2/ эндо |
С-2/ эндо для пиримидинов; С-3/ эндо для пуринов |
Конформация гликозидной связи |
Анти |
Анти |
Анти для пиримидинов; син для пуринов |

Рис. 3.8 |
Ограниченное, но возможное вращение вокруг связи 4 приводит к такому искажению кольца, при котором один из атомов пятичленного фуранозного кольца оказывается вне плоскости создаваемой четырьмя другими атомами. Такая конформация представляет собой эндо- или экзо-структуру, в зависимости от того располагается ли данный атом на той же стороне плоскости, что и С-5/ или на противоположной стороне. |
Модели трех, упомянутых форм ДНК уже были приведены выше на рис. 3.6. А-форма ДНК преобладает во многих растворителях содержащих невысокий процент воды. Эта форма ДНК все еще сохраняет структуру правосторонней двойной спирали, но диаметр этой спирали несколько больше (см. табл. 1), чем диаметр двойной спирали ДНК в В-форме и число пар нуклеотидов в одном витке составляет не 10,5 как в В-форме, а 11, при этом расстояние между парами оснований в А-ДНК по вертикали уменьшается до 2,6 Å (0,26 нм).
А-форма отличается от В-формы тем, что плоскости, в которых расположены комплементарные пары составляют с перпендикуляром к оси спирали угол, равный приблизительно 20 (табл. 1). Данные структурные особенности сопровождаются углублением большой бороздки в молекуле ДНК и, одновременно, делают малую бороздку более мелкой. Реагенты, обычно используемые для кристаллизации ДНК, обладают дегидратирующими свойствами, поэтому наиболее короткие молекулы кристаллизуются именно в А-форме.
Молекулы ДНК, существующие в Z-форме, характеризуются наиболее выраженными структурными отклонениями от классической спирали в В-фоме. Самой заметной особенностью Z-формы является ее левосторонняя спирализованность. При этом такая двухспиральная молекула становится более вытянутой, имеет меньший диаметр и содержит 12 пар оснований в одном витке спирали, а расстояние между соседними парами оснований увеличивается до 7,7 Å (0,77 нм). Название Z-форма происходит оттого, что в такой ДНК сахарофосфатный остов образует зигзагообразную линию. Z-форма существует при довольно высоких концентрациях соли, что объясняется необходимостью противостоять повышенному электростатическому отталкиванию между нуклеотидами, сжатыми из-за уменьшения диаметра двойной спирали Z-ДНК [15,16]. Установлено, что некоторые нуклеотидные последовательности имеют большую склонность к образованию левосторонней спирали, чем другие. Так, Z-форма двойной спирали найдена в полимерах или участках ДНК, образованных чередующимися динуклеотидами, состоящими из пар пурин-пиримидин.Особенно это справедливо для участков ДНК имеющих следующие многократно повторяющиеся звенья:
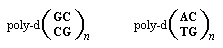
Более того, доказано, что замены остатков цитозина в таких полимерах на 5-метилцитозин в значительной степени повышают стабильность Z-ДНК в условиях близким к таковым in vivo:
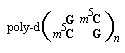
Существенным условием образования и стабилизации левосторонней спирали Z-ДНК является присутствие в ней пуриновых нуклеотидов в син-конформации, чередующихся с пиримидиновыми нуклеотидами в анти-конформации. Внешний вид Z-ДНК отличается едва заметной большой бороздкой, но узкой и глубокой малой бороздкой. Присутствие в ДНК участков в А-форме в условиях in vivo пока не доказано, хотя известно, что конформация А-ДНК близка к структуре гибридов РНК-ДНК и структуре двухспиральных РНК. Причина этого явления заключается в том, что 2/-ОН-группы в молекулах РНК препятствуют образованию классической В-формы. Напротив, относительно короткие участки Z-ДНК действительно присутствуют в этой информационной молекуле как у про-, так и эукариот. Полагают, что участки Z-ДНК играют определенную роль в регуляции экспрессии некоторых генов и принимают участие в генетической рекомбинации.
До сих пор мы рассматривали ДНК как структуру изолированную. В действительности ДНК связана с белками, которые могут оказывать значительное влияние на возможность перехода BZ. Например, ДНК, связанная с гистонами, не переходит из одной формы в другую в тех условиях, в которых это наблюдают у свободной ДНК. Поэтому одним из условий, необходимых для образования Z-формы in vivo, вероятно, является присутствие особых белков, стабилизирующих ее структуру. Большая же часть клеточной ДНК находится в В-форме с определенными модификациями параметров спирали, варьирующими локально. И только отдельные короткие участки спирали переходят в другие формы. Возможен ли переход из одной формы в другую внутри одной и той же молекулы ДНК? При определенных условиях участок, состоящий из 26-32 дуплетов d(pGpC)n. d(pCpG)n может переходить в Z-форму, тогда как участки ДНК по обе стороны от этой последовательности остаются в классической В-форме. Указанные наблюдения подтверждают идею о том, что конформационные переходы в Z-форму действительно могут иметь большое биологическое значение и происходить in vivo в специфических участках ДНК.
В настоящее время получены антитела, способные отличать Z-форму от В-формы ДНК. Эти антитела связываются с определенными хромосомами Dr. melanogaster. За реакцией связывания легко следить из-за необычного строения этих хромосом, у которых более плотные участки (диски) контрастируют с менее плотными (междисками). Участки Z-ДНК расположены в междисках. Из этого следует, что Z-форма реально существует в естественных условиях, хотя размеры индивидуальных участков Z-формы пока неизвестны.
Более детальные исследования показали, что содержание Z-ДНК in vivo сильно зависит от степени суперспиральности (см. далее) этой молекулы [19,20]. Имеются также данные о том, что последовательности, способные принимать Z-конформацию находятся в области регуляции транскрипции некоторых генов. Идентифицированы и очищены белки, специфичные к Z-ДНК. И все же, несмотря на существование Z-ДНК-сегментов в области регуляции транскрипции или по соседству с ней и белков, узнающих Z-ДНК, вопрос об участии этой формы в регуляции транскрипции пока остается открытым. Известно об одном случае участия Z-ДНК-связывающего белка в рекомбинации ДНК, но и здесь конкретный механизм влияния ДНК-связывающего белка и Z-конформации на процесс рекомбинации остается неясным.