

Рибонуклеотидредуктаза и биосинтез дезоксирибонуклеотидов
Большинство атомов углерода, проходящих через пути синтеза нуклеотидов попадает в рибонуклеозидтрифосфаты (рНТФ) - ATФ, ЦТФ, ГТФ, и УТФ. Относительно небольшая доля атомов углерода включается в синтез дезоксирибонуклеозидтрифосфатов (дНТФ). Преобладание синитеза рНТФ над синтезом дНТФ связано с тем, что большинство клеток содержит в 5-10 раз больше РНК, чем ДНК, а также ещё и потому, что рНТФ выполняют ряд специфических функций в клетке, в то время как дНТФ используются только для синтеза ДНК. Как показано на рис рНДФ ("Д" означает ди-) превращаются в соответствующие дезоксирибонуклеозиддифосфаты при участии фермента рибонуклеозиддифосфат релуктазы (также называемая рибонуклеотид редуктаза, рНДФ редуктаза или РНР). Относительно простыми являются пути синтеза дАТФ, дГТФ и дЦТФ в отличие от механизма образования дТТФ, предшественником которого служит дУДФ
Рибонуклеотид редуктаза, фермент, катализирующий синтез дНДФ из рНДФ восстанавливает гидроксил при С2 рибозы по свободно радикальному механизму.
Восстановление рибонуклеотидов требует электронов. Они в конечном счете доставляются из НАДФН+Н+ при участии тиоредоксина или глутаредоксина, как это показано на рис 8.18. Тиоредоксин широко распространен в животном и растительном мире. У животных он контролирует уровень инсулина, способствуя его восстановлению, участвует в образовании меланина (люди с высоким уровнем тиоредоксина легко загорают), способствует формированию пространственной структуры белков.
Р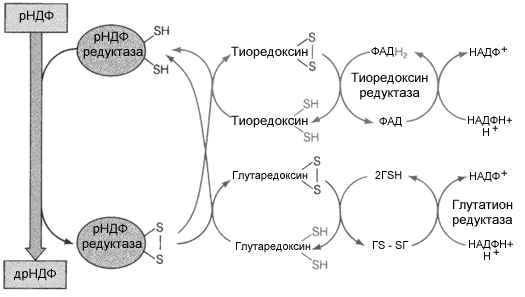 ис.8.18.Схема
реакций, катализируемых с участием рНДФ
редуктазы
ис.8.18.Схема
реакций, катализируемых с участием рНДФ
редуктазы
Биосинтез тимидиловых дезоксирибонуклеотидов
Синтез дезокситимидиловых нуклеотидов происходит иначе чем других дНТФ, которые образуются непосредственно из рибонуклеотидов под влиянием редуктазы катализирующей преобразование рибонуклеозиддифосфатов в дезоксирибонуклеозиддифосфаты (см рис.9-22).Термины тимидин и дезокситимидин (или дTTФ и ТТФ) относятся к дезоксирибонуклеотиду, потому что тимидиловый рибонуклеотид не встречается среди нормальных метаболитов. В редких случаях, где имеется тимидиловый рибонуклеотид его обычно обозначают как 'рТТФ'
Синтез тимидиловых нуклеотидов de novo начинается или с УДФ или ЦДФ и приводит к образованию дТТФ . Что касается возможной реутилизации , то образование тимидиловых нуклеотидов начинается с дезоксицитина, дезоксиуридина или дезокситимидина, которые превращаются в соответсвующие нуклеозидмонофосфаты на первом этапе при помощи соответсвующих киназ (см раздел о реутилизации нуклеотидов) Имеются несколько уровней регуляции синтеза. Например, дЦТФ ингибирует реакции реутилизации, катализируемую дезоксицитидин киназой и активизирует реакцию, катализируемую дЦМФ дезаминазой. С другой стороны, дTTФ ингибирует реакцию дезаминирования дЦМФ, а также фермент тимидин киназу.
. 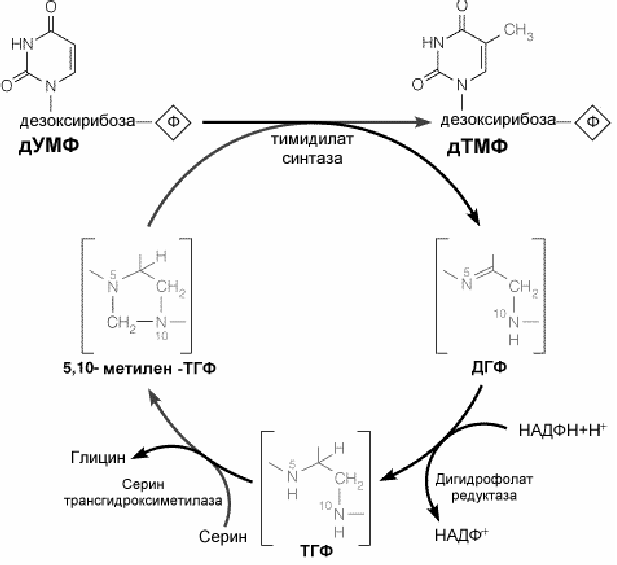
Рис.8-19.Механизм образования дезокситимидилового нуклеотида (дТМФ)
Превращение дУМФ в дТМФ сопровождается переносом одноуглеродного фрагмента от 5,10-метилентетрагидрофолата в реакции, катализируемой тимидилат синтазой.
Реакция, катализируемая тимидилатсинтазой - единственная, известная в клетке, в которой ТГФ не регенерирует. Дигидрофолатредуктаза, таким образом, играет существенную роль в окончательной регенерации 5,10 метилен-ТГФ. Этот фермент явялется мишенью при антиопухолевой химиотерапии, так как он ингибируется лекарственным средством метотрексатом.