

Сплайсинг – способ создания многообразия белков
После модификации концов иРНК наступает одна из самых сложных реакций процессинга -точное вырезание различных по длине внутренних участков (интронов ) и сшивание оставшихся, несущих смысловую нагрузку для кодируемого белка - экзонов. Совокупность реакций, происходяших при этом получила название сплайсинг.
На первый взгляд присутствие интронов в генах эукариот, казалось бы, является бессмысленной тратой клеточной энергии затрачиваемой вначале на их включение в первичный транскрипт только для того, чтобы быть удаленным позже. Однако, присутствие интронов защищают функционально активную часть генома клетки от повреждающего действия химических илифизических (лучевых) факторов. Еще одна важная функция интронов позволяет при помощи так называемого альтернативного сплайсинга увеличить генетическое разнообразие генома без увеличения числа генов. В результате изменения распределение экзонов одного транскрипта во время сплайсинга возникают различные РНК и следовательно различные белки. Альтернативный сплайсинг происходить или в определенных стадиях развития организма или в различных типах клеток.
Известны уже более 40 генов, транскрипты которых подвергаются альтернативному сплайсингу. Например, транскрипт гена кальцитонина, в результате альтернативного сплайсинга дает РНК, которая служит матрицей для синтеза кальцитонина (в щитовидной железе)или специфический белок, называемый белок, связанный с геном кальцитонина (CGRP, в мозге). Еще более сложному альтернативному сплайсингу подвергается транскрипт гена-тропомиозина. Были идентифицированы по крайней мере 8 различных тропомиозиновых иРНК, полученных из одного транскрипта (см рис)
Нарушения процессов сплайсинга может вести к различным заболеваниям. Например, талассемии связаны с дефектами генов, кодирующих–глобины. Некоторые из этих дефектов вызываются мутациями в последовательностях гена, обеспечивающих узнавание интронов
В результате происходит нарушение процессинга первичного транскрипта гена - глобина.
Еще один тип заболеваний, связанный с нарушением сплайсинга связан с образованием антител к белкам, участвующим в сплайсинге.
Р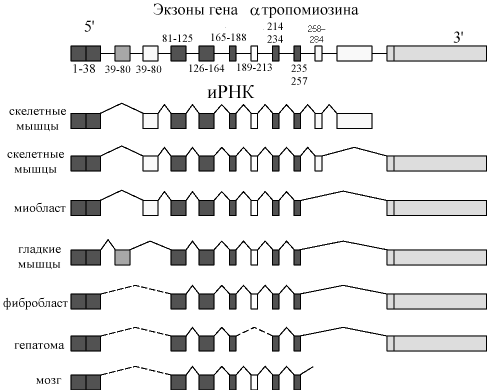 ис.8.36.Схематическое
изображение сплайсинга первичного
транскрипта гена-тропомиозина
ис.8.36.Схематическое
изображение сплайсинга первичного
транскрипта гена-тропомиозина
. Так, например, причиной системной красной волчанки - одного из тяжелых заболеваний соединительной ткани, является образование антител к комплексу белок - U1 РНК сплайсеосомы .
Процессинг продуктов рнк-полимераз I и III не похож на процессинг иРнк
Если РНК-полимераза IIкатализирует транскрипцию большинства генов, кодирующих белки, гены, которые кодируют РНК с самостоятельным функциональным значением, транскрибируются РНК-полимеразой I и РНК-полимеразой III . Обычно эти гены представлены в геноме большим числом копий, часто образующих кластеры тандемных повторов.
Первичные транскрипты генов рРНК, известные как 45S-РНК образуются РНК-полимеразой I. Их длина около 13000 нуклеотидов. Перед тем как покинуть ядро в составе собранной рибосомной частицы , молекула 45S-РНК подвергается специфическому расщеплению , в результате чего образуется по одной копии 28S-РНК (около 5000 нуклеотидов), 18S-РНК (около 2000 нуклеотидов) и 5,8S-РНК (около 160 нуклеотидов), которые собственно и являются компонентами рибосом. Общее происхождение всех трех типов рРНК из одного и того же первичного транскрипта служит гарантией того, что они образуются в равных количествах. Остальная часть этого транскрипта (около 6000 нуклеотидов) распадается в ядре. Возможно, что эти «лишние» последовательности молекулы-предшественника рРНК играют определенную роль в сборке рибосом, происходящих непосредственно по завершении синтеза 45S-РНК. Синтез предшественника рибосомных РНК происходит в ядрышке, в котором находятся гены рибосомных РНК (рДНК). рДНК представляет собой специализированные участки ( ядрышковые организаторы) нескольких хромосом. Для транскрипции рДНК in vitro, помимо самой РНК- полимеразы I, необходимы два фактора транскрипции - UBF и SL1
Транспортные РНК также синтезируются в форме больших молекул предшественников, которые затем подвергаются процессингу. Один из предшественников тРНК E. coli –представляет первичный транскрипт в 950 нуклеотидов длиной, который расщепляется рибонуклеазой P на 7 различных молекул тРНК. Рибонуклеаза P – рибонуклеопротеид, содержащий молекулу РНК в 377 нуклеотидов, которая является фактическим ферментом. Это был первый пример так называемых рибозимов – ферментов, которые представляют собой РНК, а не полипептидные цепи. После завершения расщепления первичного транскрипта тРНК происходит удаление дополнительных нуклеотидов на 5 ' и 3 ' концах и к 3' концу каждой тРНК присоединяется последовательность 5 '-ЦЦА-3',а несколько нуклеотидов основной структуры подвергаются модификации. В т-РНК были идентифицированы больше 60 различных измененных оснований .