

У эукариот – 3 рнк- полимеразы
Из клеток эукариот выделены три типа РНК-полимераз, каждая из которых ответственна за транскрипцию различных групп генов. Эти ферменты названы РНК-полимераза I, РНК-полимераза II и РНК-полимераза III (PolI,II,III). Полимеразы структурно сходны друг с другом. Молекулярная масса около 500 кД. В их состав может входить до 10 полипептидных цепей, причем некоторые из них одинаковы у разных полимераз, однако есть и уникальные, характерные для каждой полимеразы субъединицы. РНК-полимеразы эукариот и бактерий эволюционно родственны.
PolIрРНК
PolIIвсе иРНК,U1,U2,U4,U5snRNA
PolIII5SРНК, тРНК,U6snRNAи другие маленькие РНК
Как видно, только одна из полимераз, РНК-полимераза II , участвует в синтезе и-РНК, которая затем используется для синтеза белков. Кроме того эта же полимераза участвует в образовании большинства малых РНК, которые образуют snRNP.
Что касается остальных полимераз, то РНК-полимераза I синтезирует высокомолекулярную рибосомную РНК , а РНК-полимераза III - разнообразные низкомолекулярные стабильные РНК, в том числе тРНКирибосомную 5S-РНК. РНК-полимеразы эукариот нечувствительны к ингибиторам бактериальных РНК-полимераз - рифампицину и стрептолидигину. Специфическим ингибитором РНК-полимеразы II является токсин бледной поганки --аманитин, использование которого помогло в открытии разных РНК полимераз: РНК-полимераза I не чувствительна к нему; РНК-полимераза II обладает очень высокой чувствительностью ; РНК-полимераза III ингибируется только под действием высоких доз этого токсина.
Хотя синтезированные РНК-полимеразой II транскрипты составляют более половины РНК, синтезированной в результате транскрипции ДНК, большая часть РНК таких транскриптов нестабильна и, соответственно, имеет короткий промежуток жизни. Следовательно, производные от нее гетерогенные ядерные РНК (гяРНК) в клеточном ядре и цитоплазматическая мРНК составляют лишь минорную фракцию тотальной клеточной РНК
Различия в механизмах транскрипции у про и эукариот связаны также с особенностями пространственной организации этого процесса. У прокариот отсутствует ядро и поэтому синтезируемая иРНК без каких-либо изменений сразу включается в трансляцию, продолжительность использования этой РНК небольшая и следует принципу одна РНК – один белок. У эукариот эти процессы разобщены и молекулы РНК претерпевают значительные постранкрипционные изменения.
В транскрипции у прокариот важная роль принадлежит -фактору
Полный процесс инициации транскрипции и элонгации суммирован на рис.9-37. Первый шаг в транскрипции связывание РНК-полимеразы с ДНК, с последующим перемещением к инициирующему участку ДНК.
1.Поиск промотора РНК-полимеразой начинается (рис.9-37, этап 1) с неспецифического (низкое сродство) связывания холофермента с ДНК, и последующего перемещения по ДНК, без диссоциации до достижения последовательности промотора,с которым фермент связывается со значительно более высокой аффинностью. Этому способствуетфактор, потому что базовый фермент имеет одинаковое сродство к любой последовательности ДНК.
2.Первоначальное взаимодействие между холоферментом РНК-полимеразы и промоторомобразует закрытый комплекс. с константой асссоциацииKa106и 109M-1в 0.1 М.NaCl. Цепь ДНК в этом комплексе не раскручена. Взаимодействие между компонентами электростатическое, посколькуKaзависит от ионной силы. Комплекс относительно неустойчив, время его полураспада приблизительно 10 секунд.
3.РНК-полимераза раскручивает несколько пар оснований ДНК, от приблизительно -10 до -1, образуя открытый промоторный комплекс, названный так потому, что в нем цепи ДНК, открыты, или раскручены. Эта чувствительная к температуре реакция происходит с полупериодом распада приблизительно от 15 секунд до 20 минут, в зависимости от структуры промотора. Открытый промоторный комплекс чрезвычайно устойчив; его трудно разрушить высокой ионной силой (Kaболее чем 1014M-1). Затем происходит Mg2+ - зависимая изомеризация, формируя модифицированную форму открытого комплексапромоторас раскрученной областью ДНК, занимающего от-12 до +2.
После того как РНК-полимераза свяжется с промотором с образованием открытого промоторного комплекса , фермент готов начать синтез цепи РНК. Во время элонгации РНК-полимераза использует участок связывающий один нуклеозидтрифосфат.
. Он связывает любой из четырех нулеозид трифосфатов (рНТФ). Другой связывающий участок используется для инициации. Он связывает преимущественно АТФ или ГТФ, а это означает, что большинство иРНК имеют пуриновые нуклеотиды на 5 ' конце.
1. Рост цепи начинается со связывания указанного матрицей рНТФ на специфическом инициирующем участке РНКполимеразы (рис.9-37. шаг 4),
2. Следующий нуклеотид связывается со специфическим участком элонгации .
3. Нуклеофильная атака 3 ' ОН группой первого нуклеотида на (внутренний) фосфат второго нуклеотида образует первую фосфодиэфирную связь и оставляет интактной трифосфатную половину на 5 ' положении первого нуклеотида.

Рис.8-32.Основные этапы процесса транскрипции
Большинство инициирований абортивно, и завершается высвобождением олигонуклотида от 2 до 9 нуклеотидов длиной. Еще не ясно, почему это случается. После синтеза первых 10 нуклеотидов, субъединица отделяется от комплекса транскрипции, и дальнейший процесс транскрипции катализируется базовой (коровой) полимеразой (Рис, шаги 5 и 6). Хотя исубъединица покинула фермент, элонгирующий комплекс является довольно устойчивым. Транскрипция, как показаноinvitro, больше не ингибируется рифампицином. Элонгация, таким образом, продолжается до заключительного этапа.
В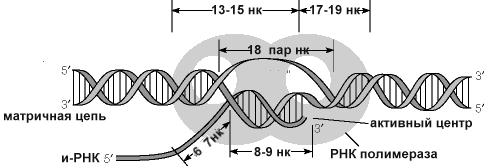 о
время элонгации (рис.8.32, шаги 5 и 6), коровый
феремент перемещается по двуцепочечной
матрице ДНК, и одновременно раскручивает
ДНК, экспонируя одноцепочечную матрицу
для образования пар с поступающими
нуклеотидами и с образованием транскрипта
(вновь синтезируемая РНК). Фермент
скручивает матрицу позади 3 ' конца
растущей цепи РНК. В модели, показанной
в рис.8-33 приблизительно 18 пар оснований ДНК
раскручены, формируя движущийся "
пузырь транскрипции. " Как только
одна пара оснований стано
о
время элонгации (рис.8.32, шаги 5 и 6), коровый
феремент перемещается по двуцепочечной
матрице ДНК, и одновременно раскручивает
ДНК, экспонируя одноцепочечную матрицу
для образования пар с поступающими
нуклеотидами и с образованием транскрипта
(вновь синтезируемая РНК). Фермент
скручивает матрицу позади 3 ' конца
растущей цепи РНК. В модели, показанной
в рис.8-33 приблизительно 18 пар оснований ДНК
раскручены, формируя движущийся "
пузырь транскрипции. " Как только
одна пара оснований стано
Рис.8.33.Схема «пузыря транскрипции» в области активного центра РНК-полимеразы
вится раскрученной перед 3 ' концом возникающей цепи РНК, одна пара оснований скручивается у хвоста молекулы полимеразы РНК. Приблизительно 8 пар оснований 3 ' конца возникающего транскрипта спарены с матричной цепью ДНК.
У E. coliтерминация транскрипции может происходить двумя способами: фактор зависимым и фактор независимыми способами. При фактор независимом способе роль терминирующего сигнала выполняют 2 симметричных богатых ГЦ сегмента, которые способны к формированию структуры шпильки. Формирование шпильки в РНК дестабилизирует ассоциацию между полимеразой и матрицей ДНК. Эта дестабилизация усиливается благодаря более слабым характером связей пар основанийAУ, которые формируются между матрицей и РНК, расположенных после шпильки. Транскрипция большинства генов уE. coliтерминируется этим способом.
Факторзависимая терминация требует узнавания последовательностей терминирующим белком rho. Этот фактор узнает и связывается с последовательностями на 3 ' конце РНК. Связывание дестабилизирует взаимодействие матрицы и полимеразы и останавливает транскрипцию.