

UnEncrypted
.pdfMeta-epidemic model
Figure 1: Scheme of the model
2Model formulation
We consider two compartmentalized environments containing each individuals of a population, subject to a transmissible disease, so that they can be either susceptible or infected, Si and Ii, i = 1, 2, respectively.
We assume that mij is the susceptibles’ migration rate from environment j to environment i, with i, j = 1, 2. Similarly, let nij denote the migration rate of infected from environment j to environment i, with i, j = 1, 2. In principle they are di erent as migrations, entailing in general an e ort, are perhaps less possible for infected, that in view of the disease, are weaker individuals. We assume the disease to be recoverable, and that the recovery rate may be influenced by the environment. Thus let ν and η denote these rates. We also assume that ecological conditions in the two environments are in principle di erent, thereby a ecting the disease propagation and its recovery. Thus, the disease transmission rate in the first environment is γ while it is β in the second one. Overall, then, we have two copies of the classical SIS model linked via migrations. A schematic representation of the situation is graphically illustrated in Figure 1.
Using mass action incidence to model disease transmission, the model thus becomes
S˙1 |
= |
−γS1I1 + νI1 − m21S1 + m12S2 |
(1) |
I˙1 |
= γS1I1 − νI1 − n21I1 + n12I2 |
|
|
S˙2 |
= −βS2I2 + ηI2 + m21S1 − m12S2 |
|
|
I˙2 |
= |
βS2I2 − ηI2 + n21I1 − n12I2 |
|
c CMMSE |
Page 123 of 1573 |
ISBN:978-84-615-5392-1 |
Marika Barengo, Isabella Iennaco, Ezio Venturino
Further, like in the classical epidemic models, [1], we make the assumption that the total population in the two enviroments has a constant value N, so that the recruitment rate of newborns matches the mortality rate, natural plus disease-related. Thus, S1 + I1 + S2 + I2 = N, and this is reflected by the fact that on summing the equations in (1), we find
S˙1 |
+ I˙1 + S˙2 + I˙2 = 0. |
We can thus express S1 as function of the remaining populations, |
||
S1 |
= N − I1 − S2 − I2 |
and therefore eliminate it from the system. |
We obtain then the |
|
simplified system |
|
|
|
|
|
I˙1 |
= |
γ (N − I1 − S2 − I2) I1 − νI1 − n21I1 + n12I2 |
(2) |
|
S˙2 |
= −βS2I2 + ηI2 + m21 (N − I1 − S2 − I2) − m12S2 |
||
|
I˙2 |
= βS2I2 − ηI2 + n21I1 − n12I2 |
|
|
2.1 Analysis of the equilibria
For the system (2) there are only two possible equilibria, in addition to the origin E(0) = (0, 0, 0), which is an equilibrium only if the total population vanishes, as consequence of
the second equation (2). These points are the disease-free point E(1) = (0, S |
(1) |
, 0) and the |
|||||
|
|
|
|
|
|
2 |
|
coexistence equilibrium, with endemic disease, E(2) = (I(2) |
, S(2) |
, I |
(2)). The former is always |
||||
|
1 |
2 |
|
2 |
|
|
|
feasible, with |
m21 |
|
|
|
|
|
|
S2(1) = NM0, M0 ≡ |
. |
|
|
|
|
||
|
|
|
|
|
|||
|
m21 + m12 |
|
|
|
|
||
To find the components of the coexistence equilibrium, we sum the second and the third equilibrium equations thereby replacing the second one, and solve the third one to get the
following algebraic system |
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||
|
|
0 = γNI1 − γI12 − γS2I1 − γI2I1 − νI1 − n21I1 + n12I2 |
|
|
||||||||||||||||
|
|
|
|
1 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
0 = n21I1 + Nm21 − m21I1 − m21S2 − m21I2 − m12S2 − n12I2 |
(3) |
|||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
I |
|
= |
|
|
(n |
|
+ η − |
βS |
)I |
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||
|
|
|
1 |
|
n21 |
12 |
|
|
2 |
|
2 |
|
|
|
|
|
|
|
||
Letting |
|
|
|
|
|
|
n21 |
|
|
|
n12 |
|
m12 |
|
|
|||||
|
|
M1 ≡ |
|
, |
M2 ≡ |
|
|
, |
M3 ≡ |
|
, |
|
||||||||
|
|
m12 + m21 |
m12 + m21 |
m12 + m21 |
|
|||||||||||||||
from the second equation we find |
|
|
|
|
|
|
|
|
|
|
||||||||||
S2 = |
Nm21 − I1m21 + n21I1 − m21I2 − n12I2 |
= M0(N − I1 − I2) + M1I1 − M2I2, |
|
|||||||||||||||||
|
|
|||||||||||||||||||
|
|
|
|
|
|
|
m12 + m21 |
|
|
|
|
|
|
|
|
|
|
|||
which immediately gives a necessary feasibility condition, namely |
|
|
||||||||||||||||||
|
|
|
|
|
|
|
M0N > (M0 − M1)I1 + (M2 + M0)I2. |
|
(4) |
|||||||||||
c CMMSE |
Page 124 of 1573 |
ISBN:978-84-615-5392-1 |
|
Meta-epidemic model |
Back substitution into (3) gives two equations in I1 e I2, |
|
EI12 + 2F I1I2 + 2GI1 + 2HI2 = 0, |
(5) |
AI22 + 2BI1I2 + 2CI2 + 2DI1 = 0, |
(6) |
where we set E ≡ −γ(M1 + M3), F ≡ 12 γ(M2 − M3), G ≡ 12 (γNM3 − n21 − ν), H ≡ 12 n12, A ≡ β(M0 + M2), B ≡ 12 β(M0 − M1), C ≡ 12 (n12 + η − βNM0) and D ≡ − 12 n21.
Equations (5) and (6) identify two conic sections, both through the origin and respectively intersecting the coordinate axes at the points (−2GE−1, 0) and (0, −2CA−1). The former conic for
|
−n12(M1 + M3) |
= γNM3 − n21 − ν, n12 = m12 |
(7) |
|||||||||
|
|
|
|
|||||||||
|
M2 − M3 |
|
|
|
|
|
|
|
|
|
||
is a hyperbola with center at (−HF −1, −(F G − EH)F −2) and asymptotes |
|
|||||||||||
|
|
H |
|
|
H |
|
F |
F G − EH |
|
|||
|
x = − |
|
, |
x + |
|
= − |
2 |
y + |
|
. |
|
|
|
F |
F |
E |
F 2 |
|
|||||||
In case n12 = m12 it becomes a parabola with axis x = − GE and vertex (−GE−1, G2(2EH)−1), with negative height there since EH = −n12γ(M1 + M3) < 0. The degenerate case
|
−n12(M1 + M3) |
|
= γNM3 − n21 − ν, |
n12 = m12 |
(8) |
||||||||||||||
|
|
M2 − M3 |
|||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
leads instead to the two straight lines |
|
|
|
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
x = − |
2F |
y, |
x + |
H |
= 0. |
|
|
|
(9) |
||||||
|
|
|
|
|
|
|
|
|
|
||||||||||
|
|
|
|
|
|
|
|
E |
|
|
F |
|
|
|
|
||||
If |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
− |
n21(M0 + M2) |
|
|
= n12 + η − βNM0, |
n21 = m21 |
(10) |
|||||||||||||
M − M |
|
||||||||||||||||||
0 |
1 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
also (6) is a hyperbola, with center at (−(BC − DA)B−2, −DB−1) and asymptotes |
|
||||||||||||||||||
|
x + |
BC − DA |
|
|
|
A |
y + |
D |
|
D |
|
||||||||
|
|
|
|
= − |
|
|
, |
y = − |
|
. |
|
||||||||
|
|
B2 |
2B |
B |
B |
|
|||||||||||||
For n21 = m21 it becomes a parabola with axis y = −CA−1 and vertex (C2(2DA)−1, −CA−1). It also has negative height at the vertex in view of the fact that 2DA = −n21β(M0 +M2) < 0. The degenerate case is obtained if
− |
n21(M0 + M2) |
= n12 + η − βNM0, n21 = m21, |
(11) |
|
|
||||
|
M − M |
1 |
|
|
0 |
|
|
||
c CMMSE |
Page 125 of 1573 |
ISBN:978-84-615-5392-1 |

Marika Barengo, Isabella Iennaco, Ezio Venturino
giving thus the two straight lines
y = − DB , x = − 2AB y. (12)
These conic sections will always intersect in the first quadrant, providing a feasible equilibrium if (4) is satisfied, but for the degenerate cases for which HF > 0 and DB > 0 (or equivalently F E < 0 and AB < 0). In the degenerate cases, we can explicitly write the equilibrium:
i) − |
H |
> 0, |
− |
D |
> 0 imply E |
(2) |
= |
|
|
−n12 |
, |
|
n21 |
, |
n12(M0 |
−M1) |
+ |
n12 |
+ |
η |
; |
|
|
||||||||||
F |
B |
|
|
|
γ(M2−M3) |
β(M0−M1) |
γ(M2 |
−M3) |
|
β |
β |
|
|
||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||
|
H |
|
|
D |
|
(2) |
= |
|
|
−n12 |
|
|
|
(M0−M1)n12 |
|
|
|
|
|
|
(M0−M1)(n12−n21) |
|
|||||||||||
ii) − F |
> 0, |
− B |
< 0 imply E |
|
|
|
|
|
, |
|
|
|
|
|
, NM0 + |
|
|
|
; |
||||||||||||||
|
|
|
γ(M2−M3) |
|
γ(M0+M2)(M2−M3) |
|
|
γ(M2−M3) |
|||||||||||||||||||||||||
|
H |
< 0, |
|
D |
> 0 imply E |
(2) |
= |
|
|
(M2−M3)n21 |
|
, |
|
n21 |
, |
(M2−M3)n21 |
|
n12 |
η |
|
|||||||||||||
iii) − F |
− B |
|
|
|
|
|
|
+ |
|
|
+ β . |
|
|||||||||||||||||||||
|
|
|
(M1+M3)(M0−M1) |
β(M0−M1) |
(M1+M3)β |
|
β |
|
|||||||||||||||||||||||||
From the equilibria that we just found, we observe that it is not possible that the disease becomes a pandemic in the environment, i.e. it invades the whole population and the susceptibles totally disappear. This represents a good result from the epidemiological point of view.
2.2Stability
For the stability analysis we write the Jacobian J = (Jik), i, k = 1, 2, 3 of the system (2):
−2γI1(i) |
+ γ(N − I2(i) − S2(i)) − ν − n21 |
−γI1(i) |
− m12 |
−γI1(i) + n12 |
|
(13) |
|
−m21 |
−βI2(i) − m21 |
−βS2(i) + η − m21 |
|||
|
|
|
|
|
|
|
|
n21 |
βI2(i) |
|
βS2(i) − η − n12 |
|
|
For the origin E(0), one eigenvalue can be immediately evaluated, −(m12 + m21) < 0, the remaining ones are the roots of a quadratic stemming from a suitable 2 × 2 submatrix
J |
|
he Routh-Hurwitz conditions reduce to |
|
0 |
of J. T |
−tr(J0) = ν + η + n21 + n12 > 0, |
det(J0) = ην + νn12 + ηn21 > 0, |
|
|
|
|
so that E(0) is always stable. Recall again, compare the discussion at the beginning of Section 2.1, that this means that the whole population is wiped out, including S1 which do not explicitly appear in the model. This result is not good from the environmental point of view, since the population disappears completely, but it should be expected. In fact, if the population drops it will never be able to recover as no such specific mechanisms are present in the model.
c CMMSE |
Page 126 of 1573 |
ISBN:978-84-615-5392-1 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Meta-epidemic model |
|
||||||||
For E(1) the eigenvalues of (13) are λ1 = −m21 −m12 < 0 and the roots of the quadratic |
|
||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
λ2 − λ(γN − γNM0 − ν − n21 + βNM0 − η − n12) |
|
|
|
|
|
|
|
|
|
||||||||||||||||||||||||||||
|
|
|
|
|
|
+(γN2βM − ηγN − n |
12 |
γN − γN2βM2 + γNM η |
|
|
|
|
|
|
|
|
|
||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
0 |
|
|
|
|
|
|
|
|
|
|
|
|
|
0 |
|
|
|
0 |
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
+γNM0n12 − νβNM0 + νη + νn12 − βNM0n21 + ηn21) = 0, |
|
|
|
|
|||||||||||||||||||||||||||||||||
for which the Routh-Hurwitz criterion gives the stability conditions |
|
|
|
|
|
|
|
|
|
||||||||||||||||||||||||||||||||||
NM0(γ − β) > γN − ν − n21 − η − n12, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
(14) |
|
||||||||||||||||||||||
NM0[γNβ(1 − M0) + γ(η + n12) − β(ν + n21)] > (η + n12)(γN − ν) − ηn21. |
|
|
|||||||||||||||||||||||||||||||||||||||||
|
|
|
|||||||||||||||||||||||||||||||||||||||||
A Hopf bifurcation should arise when the value of the parameter M0 crosses the critical |
|
||||||||||||||||||||||||||||||||||||||||||
value |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
γN − ν − n21 |
− η − n12 |
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
M |
† |
≡ |
. |
|
|
|
|
|
|
|
|
|
(15) |
|
||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
0 |
|
|
|
|
|
N(γ − |
β) |
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||
Assuming at first γ > β, a feasible bifurcation arises only if the total population size is large |
|
||||||||||||||||||||||||||||||||||||||||||
enough, namely for |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||
|
|
|
|
|
|
|
1 |
|
(ν + n21 |
+ η + n12) > N > |
1 |
(ν + n21 + η + n12). |
|
|
|
|
|
|
|
||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||||||||||
|
|
|
|
|
|
|
|
β |
|
γ |
|
|
|
|
|
|
|
||||||||||||||||||||||||||
Conversely, the result also holds for γ < β if the above inequalities are all reversed. In |
|
||||||||||||||||||||||||||||||||||||||||||
spite of this theoretical result, our simulation have not been able to reveal limit cycles. |
|
||||||||||||||||||||||||||||||||||||||||||
We therefore conjecture that there might be some incompatibility among the above given |
|
||||||||||||||||||||||||||||||||||||||||||
conditions. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
For the coexistence equilibrium E(2), the caracteristic equation is the cubic λ3 + a2λ2 + |
|
||||||||||||||||||||||||||||||||||||||||||
a1λ + a0 = 0, for which the Routh-Hurwitz stability conditions are a0 > 0, a2 > 0 and |
|
||||||||||||||||||||||||||||||||||||||||||
a2a1 > a0. Explicitly, in terms of the Jacobian components, they become |
|
|
|
|
|
|
|||||||||||||||||||||||||||||||||||||
J |
(2)J(2)J(2) |
< βI2J |
(2)J(2) |
+ γI1m21J |
(2) + n21γI1J |
(2) + m21βI2J(2) |
+ n21J(2)J |
(2), |
|
||||||||||||||||||||||||||||||||||
|
22 |
33 |
11 |
|
|
|
|
11 |
|
23 |
|
|
|
|
|
|
|
33 |
|
|
|
|
|
23 |
|
|
|
|
13 |
|
|
22 |
|
13 |
|
||||||||
J |
(2) |
+ J |
(2) |
+ J |
(2) |
|
< 0, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
(16) |
|||||
|
11 |
|
|
22 |
|
|
33 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
βI2[J(2)J(2) |
+ J(2)J(2) |
− J(2)m21] + γI1m21[J(2) |
+ J(2) |
] + n21J(2) |
[J |
(2) |
+ J(2) |
] < n21J(2) |
γI1 |
||||||||||||||||||||||||||||||||||
|
|
23 |
22 |
|
23 |
|
33 |
|
|
13 |
|
|
|
|
|
|
|
|
|
11 |
|
22 |
|
|
|
13 |
|
|
11 |
|
33 |
|
|
23 |
|
||||||||
+2J |
(2) |
J |
(2) |
J |
(2) |
+ (J |
(2) |
|
2 |
(2) |
+ J |
(2) |
) + (J |
(2) |
|
2 |
|
(2) |
+ J |
(2) |
) + (J |
(2) |
2 |
(2) |
+ J |
(2) |
). |
|
|||||||||||||||
22 |
33 |
11 |
11 |
) (J |
22 |
|
33 |
22 |
) (J |
|
|
|
|
33 |
|
) (J |
22 |
11 |
|
|
|||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
11 |
33 |
|
|
|
|
|
|
|
|
|
|||||||||||||
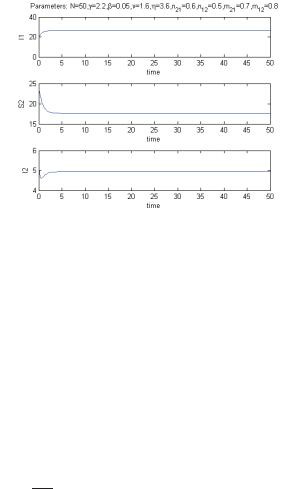
For the parameters N = 50, γ = 2.2, β = 0.05, ν = 1.6, η = 3.6, n21 = 0.6, n12 = 0.5, m21 = 0.7, m12 = 0.8, we find the coexistence equilibrium indeed at a stable state, see Figure 2.
3The case of migration only in one direction
We assume now that migration from patch 2 into patch 1 is not possible, m12 = n12 = 0,
for which M |
|
M |
|
M |
E(k), k = 0, . . . , 3. |
0 |
= 1, |
2 |
= |
|
3 = 0. The equilibria are here denoted by |
c CMMSE |
Page 127 of 1573 |
ISBN:978-84-615-5392-1 |
Marika Barengo, Isabella Iennaco, Ezio Venturino
Figure 2: Coexistence at a stable state.
For E(0) ≡ E(0) = (0, 0, 0), the results of the stability analysis still hold, together with
|
ecological consequences. |
|
|
|
|
|
|
|
|
|
their |
|
|
|
|
(1) |
|
|
|
|
|
|
|
|
|
|
|
= N, always feasible. |
||||
For equilibrium E(1), we now have S |
2 |
|||||||||
|
|
|
|
arises, namely E(3) |
with nonvanishing components |
|||||
For this model an extra equilibrium |
|
|
|
|
||||||
|
|
|
β |
|
|
|
|
η |
|
|
|
|
|
|
|
β |
|
||||
|
(3) |
|
η |
|
|
(3) |
|
|
||
|
S2 |
= |
|
, |
|
I2 |
= N − |
|
. |
|
which is feasible for |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
η < βN. |
|
|
|
(17) |
|
For the coexistence equilibrium (2) in this case recall that M0 = 1, M2 = M3 = 0.
E
Thus S2 = N − I1 − I2 + mn2121 I1 > 0. The conic (5) becomes now just the pair of straight lines
|
|
|
|
|
|
|
I1 = 0, |
|
I1 |
= − |
(ν + n21)m21 |
< 0. |
|
|
|||||
|
|
|
|
|
|
|
γn21 |
|
|
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
Therefore it intersects the second conic (6), which now is the hyperbola |
|
||||||||||||||||||
|
|
|
|
|
β |
+ |
β |
|
β |
I1I2 + |
η − βN |
|
|
|
|||||
|
|
|
|
|
|
I22 |
|
− |
|
|
|
|
I2 − I1 |
= 0, |
|
||||
|
|
|
|
|
n21 |
n21 |
m21 |
n21 |
|
||||||||||
only at the origin and at the point M(0, N − η ), giving back the equilibria E(1) |
and E(3). |
||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
β |
|
|
|
|
|
Thus in |
this case no coexistence equilibrium is possible. |
|
|||||||||||||||||
|
(1) |
the eigenvalues are λ1 |
|
|
|
|
|
|
|
|
|||||||||
|
At E |
|
= −ν − n21 < 0, λ2 = −m21 < 0, λ3 = βN − η, giving |
||||||||||||||||
|
stability condition |
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
the |
|
|
|
|
|
|
|
|
|
|
|
η > βN. |
|
|
|
|
(18) |
||
c CMMSE |
Page 128 of 1573 |
ISBN:978-84-615-5392-1 |
Meta-epidemic model
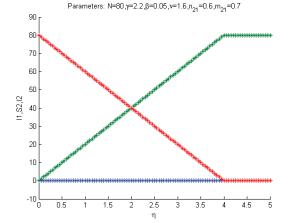
Figure 3: Transcritical bifurcation as function of the parameter η, for the case with the migrations from environment 2 to environment 1 forbidden.
Thus in this case the disease is eradicated and the whole healthy population settles only in patch 2.
For E(3) the eigenvalues are λ1 = −m21 < 0, λ2 = −ν − n21 < 0, λ3 = −βN + η. Thus, |
||||||||
follows for η < βN, which is always satisfied in view of the feasibility condition |
||||||||
stability |
(3) |
is always stable. |
|
|
|
|
|
|
(17). Hence, when feasible, E |
|
|
|
|
|
|
|
|
a transcritical bifurcation arises when E(3) |
and E |
(1) |
collide, for |
|||||
These results show that |
|
|
|
|||||
instance as a result in a change in the parameter η crossing the |
critical value η† ≡ |
βN, or, |
||||||
† |
−1 |
|
|
|||||
alternatively, if the total population crosses the critical value N |
≡ ηβ |
|
, see Figure 3, for |
|||||
the parameter values N = 80, γ = 2.2, β = 0.05, γ = 1.6, n21 = 0.6, m21 = 0.7.
Thus if the ratio of disease recovery over the disease incidence in the second environment
is lower than the total population , see (18), then (1) is the only stable equilibrium, so
N E
that the disease is eradicated and the whole population settles in the second enviroment. This is shown in Figure 4 for the parameter values N = 50, γ = 2.2, β = 0.05, γ = 1.6, η = 3.6, n21 = 0.6, m21 = 0.7.
On the contrary, if the above condition is not satisfied, the disease becomes endemic and the whole population, susceptibles and infected, still settles in the second environment,
since in this case the equilibrium (3) becomes feasible, (17) and is stable. See Figure 5
E
for a graphical description with the parameter values N = 80, γ = 2.2, β = 0.05, γ = 1.6, η = 3.6, n21 = 0.6, m21 = 0.7.
c CMMSE |
Page 129 of 1573 |
ISBN:978-84-615-5392-1 |
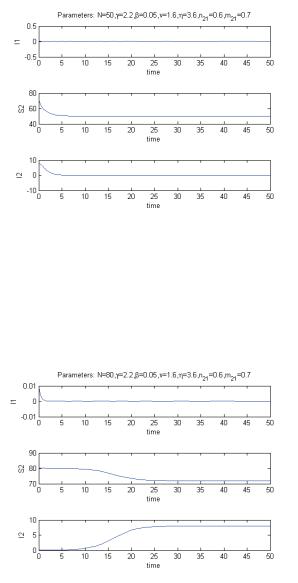
Marika Barengo, Isabella Iennaco, Ezio Venturino
Figure 4: Equilibrium (1) for the case with the migrations from environment 2 to environ-
E
ment 1 forbidden.
Figure 5: Equilibrium (3) for the case with the migrations from environment 2 to environ-
E
ment 1 forbidden.
c CMMSE |
Page 130 of 1573 |
ISBN:978-84-615-5392-1 |
Meta-epidemic model
4Infected do not migrate
In this situation, let us denote the equilibria as E(k), for k = 0, . . . , 4. Here n12 = n21 = 0, so that M1 = M2 = 0. Again at the origin the conclusions of the original system (2) still hold. In particular we then find E(1) ≡ E(1), E(3) with nonzero components
S2(3) = |
η |
, I2(3) |
= N − |
|
η |
, |
|
|||
|
|
|
|
|||||||
|
|
β |
|
βM0 |
||||||
and a new equilibrium, E(4), whose nonvanishing components are |
||||||||||
I1(4) = |
γNM3 − ν |
, |
S2(4) = |
|
M0ν |
. |
||||
|
|
|||||||||
|
|
γM3 |
|
|
|
γM3 |
||||
E(3) is feasible for |
|
|
|
|
|
|
|
|
|
|
|
|
|
βNM0 > η, |
|
(19) |
|||||
while the feasibility condition for E(4) is |
|
|
|
|
|
|
||||
|
|
|
γNM3 > ν. |
|
(20) |
|||||
For the coexistence equilibrium E(2) we have the following considerations. The two conic sections become now both degenerate: in the first case we have I1 = 0 and I2 = −I1 − (ν +γM0N)(γM3)−1, for the second one instead I2 = 0 and I2 = −N−1I1 +1−η(βNM0)−1. No intersections of these lines in the interior of the first quadrant are therefore possible, hence no coexistence equilibrium exists in this case.
The eigenvalues of the Jacobian at E(1) are −m12 − m21 < 0 and the pair βNM0 − η,
γNM3 − ν giving the stability condition |
|
|
|
|
|
|
|
|
|
|
||||
N < min ! |
|
η |
|
|
, |
|
|
ν |
|
, ". |
(21) |
|||
βM |
0 |
γM |
3 |
|||||||||||
|
|
|
|
|
−1 |
|
|
|
|
|
|
|
||
At E(3) the eigenvalues are γηM3 βM0 |
|
|
− ν and the roots of a quadratic, for which |
|||||||||||
the Routh-Hurwitz criterion in view of the feasibility condition (19) reduces to |
|
|||||||||||||
|
M3 |
< |
βν |
. |
|
|
(22) |
|||||||
|
|
|
|
|
|
|
|
|
||||||
|
M0 |
|
γη |
|
|
|
||||||||
Finally, at E(4) the eigenvalues are βνM0(γM3)−1 − η giving |
|
|||||||||||||
|
|
M3 |
> |
|
βν |
|
|
|
(23) |
|||||
|
|
M0 |
|
γη |
|
|
||||||||
|
|
|
|
|
|
|
||||||||
c CMMSE |
Page 131 of 1573 |
ISBN:978-84-615-5392-1 |
Marika Barengo, Isabella Iennaco, Ezio Venturino
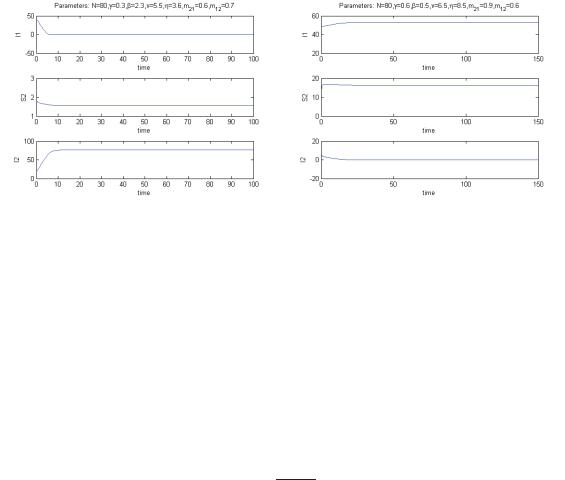
Figure 6: Left: stability of the equilibrium E(3). Right: stability of the equilibrium E(4).
and the roots of the quadratic |
|
|
|
+ m12 γN − |
|
|
|
|||||
λ2 − λ − m21 − m12 − γN + |
ν |
ν |
|
= 0 |
||||||||
M3 |
M3 |
|||||||||||
for which the Routh-Hurwitz conditions are |
|
γN − |
|
|
|
|
|
|
||||
m12 + m21 + γN − |
ν |
|
> 0 |
m12 |
ν |
|
|
> 0. |
(24) |
|||
M |
3 |
M |
3 |
|
||||||||
But the inequalities (24) hold unconditionally when the equilibrium is feasible, in view of (20), so that stability reduces to just condition (23).
Again, there is a transcritical bifurcation when E(3) and E(4) exchange their stability. This is influenced by the value of the parameter
ρ≡ βνM0 γηM3
being smaller or larger than 1. Further, when E(1) is stable, it is the only feasible equilibrium, while when either E(3) or E(4) are feasible, E(1) is always infeasible.
Thus in this case the disease gets eradicated from the ecosystem, if the system settles to equilibrium E(1). Else at E(3) it is eradicated only in environment 1, see Figure 6 left, or finally at E(4) it is eradicated only in environment 2, see Figure 6 right. No other possibilities exist, when the ecosystem thrives.
5Conclusions
We now discuss briefly some consequences that can be drawn from the above analyses. Recall that all these systems are fragile, in the sense that the whole population can vanish,
c CMMSE |
Page 132 of 1573 |
ISBN:978-84-615-5392-1 |