

Пентозофосфатный цикл
Окислительный этап образования пентоз и неокислительный этап (путь возвращения пентоз в гексозы) составляют вместе циклический процесс.
Такой процесс можно описать общим уравнением:
6 Глюкозо-6-фосфат + 12 NADP+ + 2 Н2О → 5 Глюкозо-6-фосфат + 12 NADPH +12 Н+ + 6 СO2.
Это означает, что из 6 молекул глюкозы образуются 6 молекул рибулозо-5-фосфат (пентозы) и 6 молекул СО2. Ферменты неокислительной фазы превращают 6 молекул рибулозо-5-фосфат в 5 молекул глюкозы (гексозы). При последовательном проведении этих реакций единственным полезным продуктом является NADPH, образующийся в окислительной фазе пентозофосфатного пути. Такой процесс назьюают пентозофосфатным циклом.

Пентозофосфатный цикл в жировой ткани.
Протекание пентозофосфатного цикла позволяет клеткам продуцировать NADPH, необходимый для синтеза жиров, не накапливая пентозы.
Энергия, выделяющаяся при распаде глюкозы, трансформируется в энергию высокоэнергетического донора водорода - NADPH. Гидрированный NADPH служит источником водорода для восстановительных синтезов, а энергия NADPH преобразуется и сохраняется во вновь синтезированных веществах, например жирных кислотах, высвобождается при их катаболизме и используется клетками.
Дефект глюкозо-6-фосфат дегидрогеназы в эритроцитах
Неферментативное окисление гемоглобина (Fe2+) в метгемоглобин (Fe3+) приводит к одноэлектронному восстановлению кислорода и появлению реакционно-способного анион-радикала - супероксида 02-, который служит предшественником других активных форм кислорода: пероксида водорода Н2О2 и гидроксильного радикала ОН-. Активные формы кислорода являются сильнейшими окислителями и поэтому способны вызывать серьёзные повреждения молекул ДНК, белков, ненасыщенных липидов.
В эритроцитах, как и в большинстве клеток, присутствует тиолсодержащий трипептид - глутатион (γ-глутамил-цистенил-глицин). Восстановленная форма глутатиона (Г-SH) содержит SH-группу, которая может служить донором электронов в реакциях восстановления. Под действием фермента глутатионпероксидазы восстановленный глутатион превращает молекулу пероксида водорода в молекулу воды, а сам переходит в окисленное состояние (Г-88-Г). Регенерацию восстановленного глутатиона обеспечивает глутатионредуктаза, используя в качестве донора водорода гидрированный NADPH. Для эритроцитов единственным источником получения NADPH служит пентозофосфатный путь, для других тканей существует альтернативный способ - при участии NADH-зависимой малатде-гидрогеназы (малик-фермент).
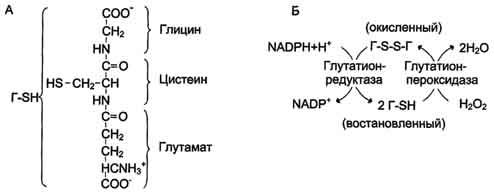
Восстановление глутатиона под действием глутатионредуктазы. А - строение глутатиона; Б - восстановление глутатиона.
Взаимодействие восстановленного глутатиона с пероксидом водорода в эритроцитах предохраняет цистеиновые остатки в протомерах гемоглобина от окисления. При генетическом дефекте глюкозо-6-фосфатдегидрогеназы концентрация восстановленного кофермента NADPH уменьшается, в результате чего резко снижается концентрация восстановленного глутатиона, а в клетке, соответственно, увеличивается количество активных форм кислорода. В этом случае окисление SH-групп молекул гемоглобина в эритроцитах приводит к образованию перекрёстных дисульфидных связей и агрегации протомеров гемоглобина с формированием телец Хайнца. В присутствии телец Хайнца пластичность мембраны нарушается, и она теряет способность к деформации при прохождении эритроцитов через капилляры. Это вызывает нарушение целостности мембраны, что приводит к гемолизу эритроцитов. Некоторые лекарственные вещества, например антималярийный препарат примахин, сульфаниламиды, также снижают способность
эритроцитов бороться с активными формами кислорода.