

Регуляция активности гликогенфосфорилазы и глигогенсинтазы
Действие этих гормонов в конечном счете сводится к изменению скорости реакций, катализируемых ключевыми ферментами метаболических путей обмена гликогена – гликогенсинтазой и гликогенфосфорилазой, активность которых регулируется аллостерически и фосфорилированием/ десфорилированием.
Гликогенфосфорилаза существует в 2 формах:
1) Фосфорилированная - активная (форма а); 2) дефосфорилированная - неактивная (форма в).
Фосфорилирование осуществляется путём переноса фосфатного остатка с АТФ на гидроксильную группу одного из сериновых остатков фермента. Следствие этого - конформационные изменения молекулы фермента и его активация.
Взаимопревращения 2 форм гликогенфосфорилазы обеспечиваются действием ферментов киназы фосфорилазы и фосфопротеинфосфатазы (фермент, структурно связанный с молекулами гликогена). В свою очередь, активность киназы фосфорилазы и фосфопротеинфосфатазы также регулируется путём фосфорилирования и дефосфорилирования.
Активация киназы фосфорилазы происходит под действием протеинкиназы А - ПКА (цАМФ-зависимой). цАМФ сначала активирует протеинкиназу А, которая фосфорилирует киназу фосфорилазы, переводя её в активное состояние, а та, в свою очередь, фосфорилирует гликогенфосфорилазу. Синтез цАМФ стимулируется адреналином и глюкагоном.
Активация фосфопротеинфосфатазы происходит в результате реакции фосфорилирования, катализируемой специфической протеинкиназой, которая, в свою очередь, активируется инсулином посредством каскада реакций с участием других белков и ферментов. Активируемая инсулином протеинкиназа фосфорилирует и тем самым активирует фосфопротеинфосфатазу. Активная фосфопротеинфосфатаза дефосфорилирует и, следовательно, инактивирует киназу фосфорилазы и гликогенфосфорилазу.

Влияние инсулина на активность гликогенсинтазы и киназы фосфорилазы. ФП-фосфатаза (ГР) - фосфопроте-инфосфатаза гранул гликогена. ПК (pp90S6) - протеинкиназа, активируемая инсулином.
Активность гликогенсинтазы также изменяется в результате фосфорилирования и дефосфорилирования. Однако есть существенные различия в регуляции гликогенфосфорилазы и гликогенсинтазы:
-
фосфорилирование гликогенсинтазы катализирует ПК А и вызывает её инактивацию;
-
дефосфорилирование гликогенсинтазы под действием фосфопротеинфосфатазы, наоборот, её активирует.
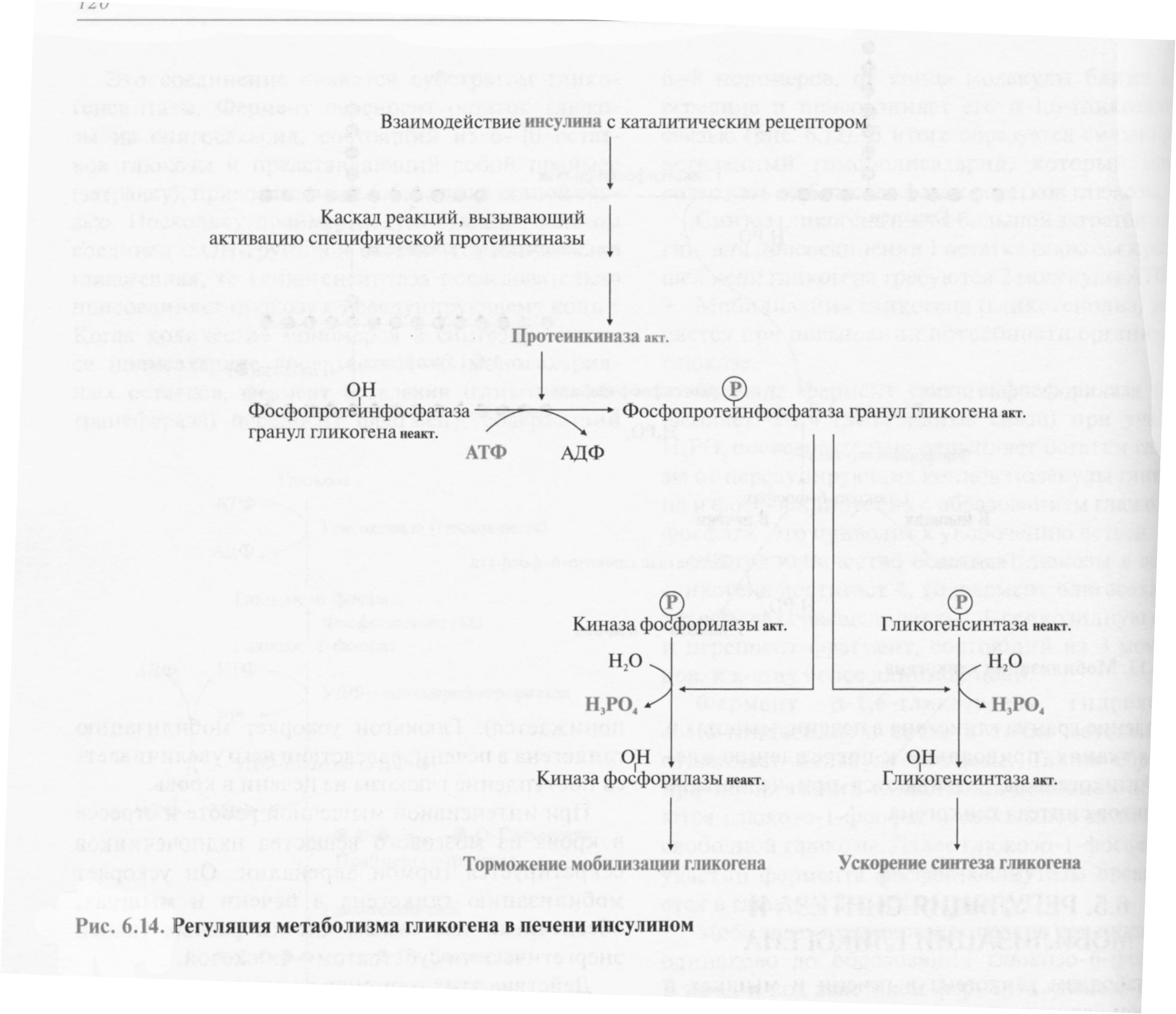
Регуляция метаболизма гликогена в печени
Повышение уровня глюкозы в крови стимулирует синтез и секрецию β-клетками поджелудочной железы гормона инсулина. Инсулин передает сигнал в клетку через мембранный каталитический рецептор - тирозиновую протеинкиназу. Взаимодействие рецептора с гормоном инициирует ряд последовательных реакций, приводящих к активации фосфопротеинфосфатазы гранул гликогена. Этот фермент дефосфорилирует гликогенсинтазу и гликогенфосфорилазу, в результате чего гликогенсинтаза активируется, а гликогенфосфорилаза становится неактивной.
Таким образом, в печени ускоряется синтез гликогена и тормозится его распад.
При голодании снижение уровня глюкозы в крови является сигналом для синтеза и секреции α-клетками поджелудочной железы глюкагона. Гормон передает сигнал в клетки через аденилатциклазную систему. Это приводит к активации проттеинкиназы А, которая фосфорилирует гликогенсинтазу и киназу фосфорилазы. В результате фосфорилирования гликогенсинтаза инактивируется и синтез гликогена тормозится, а киназа фосфорилазы становится активной и фосфорилирует гликогенфосфорилазу, которая становится активной. Активная гликогенфосфорилаза ускоряет мобилизацию гликогена в печени.

1 - глюкагон и адреналин взаимодействуют со специфическими мембранными рецепторами. Комплекс гормон-рецептор влияет на конформацию G-белка, вызывая диссоциацию его на протомеры и замену в α-субъединице ГДФ на ГТФ;
2 - α-субъединица, связанная с ГТФ, активирует аденилатциклазу, катализирующую синтез цАМФ из АТФ;
3 - в присутствии цАМФ протеинкиназа А обратимо диссоциирует, освобождая обладающие каталитической активностью субъединицы С;
4 - протеинкиназа А фосфорилирует и активирует киназу фосфорилазы;
5 - киназа фосфорилазы фосфорилирует гликогенфосфорилазу, переводя её в активную форму;
6 - протеинкиназа А фосфорилирует также гликогенсинтазу, переводя её в неактивное состояние;
7 - в результате ингибирования гликогенсинтазы и активации гликогенфосфорилазы гликоген включается в процесс распада;
8 - фосфодиэстераза катализирует распад цАМФ и тем самым прерывает действие гормонального сигнала. Комплекс α-субъединица-ГТФ затем распадается.
При интенсивной физическеой работе и стрессе в крови повышается концентрация адреналина. В печени есть два типа мембранных рецепторов адреналина. Эффект адреналина в печени обусловлен фосфорилированием и активацией гликогенфосфорилазы. Адреналин имеет сходный с глюкагоном механизм действия. Но возможно включение и другой эффекторной системы передачи сигнала в клетку печени.

Регуляция синтеза и распада гликогена в печени адреналином и Са2+.
ФИФ2- фосфатидилинозитолбисфосфат; ИФ3- инозитол-1,4,5-трифосфат; ДАГ - диацилглицерол; ЭР - эндоплазматический ретикулум; ФС - фосфодитилсерин.
1 - взаимодействие адреналина с α1-рецептором трансформирует сигнал через активацию G-белка на фосфолипазу С, переводя её в активное состояние;
2 - фосфолипаза С гидролизует ФИФ2 на ИФ3 и ДАГ;
3 - ИФ3 активирует мобилизацию Са2+ из ЭР;
4 - Са2+, ДАГ и фосфодитилсерин активируют протеинкиназу С. Протеинкиназа С фосфорилирует гликогенсинтазу, переводя её в неактивное состояние;
5 - комплекс 4Са2+- кальмодулин активирует киназу фосфорилазы и кальмодулин-зависимые протеинкиназы;
6 - киназа фосфорилазы фосфорилирует гликогенфосфорилазу и тем самым её активирует;
7 - активные формы трёх ферментов (кальмодулинзависимая протеинкиназа, киназа фосфорилазы и протеинкиназа С) фосфорилируют гликогенсинтазу в различных центрах, переводя её в неактивное состояние.
Какая система передачи сигнала в клетку будет использована, зависит от типа рецепторов, с которыми взаимодействует адреналин. Так, взаимодействие адреналина с β2-рецепторами клеток печени приводит в действие аденилатциклазную систему. Взаимодействие же адреналина с α1-рецепторами "включают" инозитолфосфатный механизм трансмембранной передачи гормонального сигнала. Результат действия обеих систем - фосфорилирование ключевых ферментов и переключение процессов с синтеза гликогена на его распад. Следует отметить, что тип рецепторов, который в наибольшей степени вовлекается в ответ клетки на адреналин, зависит от концентрации его в крови.
В период пищеварения преобладает влияние инсулина, так как инсулин-глюкагоновый индекс в этом случае повышается. В целом инсулин влияет на обмен гликогена противоположно глюкагону. Инсулин снижает концентрацию глюкозы в крови в период пищеварения, действуя на метаболизм печени следующим образом:
-
снижает уровень цАМФ в клетках и тем самым активируя протеинкиназу В. Протеинкиназа В, в свою очередь, фосфорилирует и активирует фосфодиэстеразу цАМФ - фермент, гидролизующий цАМФ с образованием АМФ;
-
активирует фосфопротеинфосфатазу гранул гликогена, которая дефосфорилирует гликогенсинтазу и таким образом её активирует. Кроме того, фосфопротеинфосфатаза дефосфорилирует и, следовательно, инактивирует киназу фосфорилазы и гликогенфосфорилазу;
-
индуцирует синтез глюкокиназы, тем самым ускоряя фосфорилирование глюкозы в клетке.