













Глава 6
Распространение и передача нервных импульсов
Нервные системы животных – это, несомненно, самые сложно организованные структуры, когда–либо существовавшие на Земле. Нервная система человека, например, состоит из 1010–1011 нейронов, а кроме того, из такого же или даже большего числа невозбудимых «опорных» клеток–сателлитов, которые называются глиальными клетками, или нейроглией. В ходе развития все эти клетки удивительным образом самоорганизуются во взаимодействующие цепи – нервные сети, из которых и состоит нервная система. Большинство нейробиологов считают, что в основе многих удивительных функций нервной системы, в том числе управления движениями, восприятия, научения и сознания, лежат физические или химические процессы, протекающие в нервных структурах. Стремление понять, каким образом деятельность нервной системы обеспечивает восприятие, научение, сознание и все другие проявления поведения животных, – это, несомненно, один из самых дерзких вызовов, который был брошен Природе человеком. Пока мы можем лишь гадать о том, удастся ли когда–либо свести сложные психические процессы к физическим и химическим закономерностям.
Несмотря на сложнейшую организацию нервной системы, многое о фундаментальных механизмах ее деятельности уже известно. Работа нервной системы в значительной степени зависит от электрической активности возбудимых нервных клеток– нейронов, число которых у животных, как мы уже отмечали, огромно. К счастью для нейробиологов, это не означает, что существует столь же большое разнообразие вырабатываемых ими сигналов. Иными словами, сложность нервной системы определяется не большим разнообразием нервных сигналов, участвующих в передаче информации, а огромным количеством и сложностью взаимодействий между нейронами. На самом деле «палитра» нервных сигналов, которые нейроны передают друг другу, весьма небогата. Более того, в основе всех этих сигналов лежат те механизмы, которые мы уже в значительной степени рассмотрели в гл. 5. К этим механизмам относятся: 1) запасание электрохимически энергии в виде концентрационных градиентов ионов по разные стороны клеточной мембраны; 2) высвобождение этой энергии в виде ионных токов, пpoникающих через селективные мембранные каналы специальными воротными механизмами; 3) пассивные электрические свойства мембраны (т. е. электрическая проводимость и емкость).
6.1. Нервные клетки
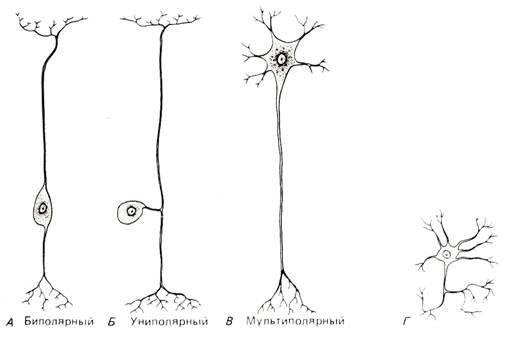
Нервные клетки различаются как по форме, так и по размерам (рис. 6–1). Их можно подразделять несколькими способами в зависимости от используемых морфологических особенностей. Один из важных критериев для такого разделения – это наличие или отсутствие у клеткиаксона (длинного нервна волокна). Знаменитый анатом Камильо Гольджи назвал нейроны с аксонами клетками типа I, а без аксонов – клетками типа II. Последние образуют связи лишь в пределах локальных нейронных контуров с непосредственно прилегающими к ним клетками.
|
|
|
Рис. 6.1. Четыре морфологические разновидности нейронов. В пределах каждой разновидности в свою очередь наблюдается значительное разнообразие. Нейроны А, Б, и В относятся к нейронам типа I по Гольджи, а нейрон Г (без аксона) к нейронам типа II. (Montagna, 1959.)
|

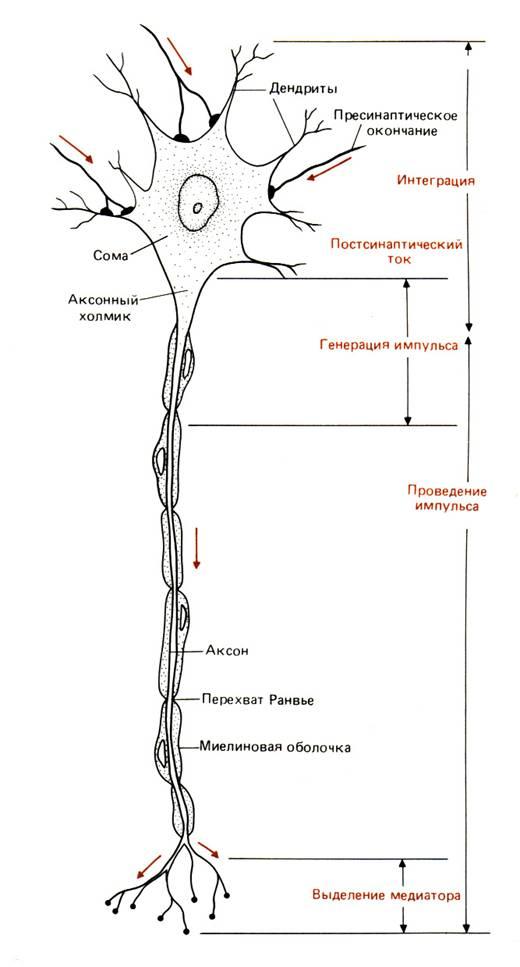
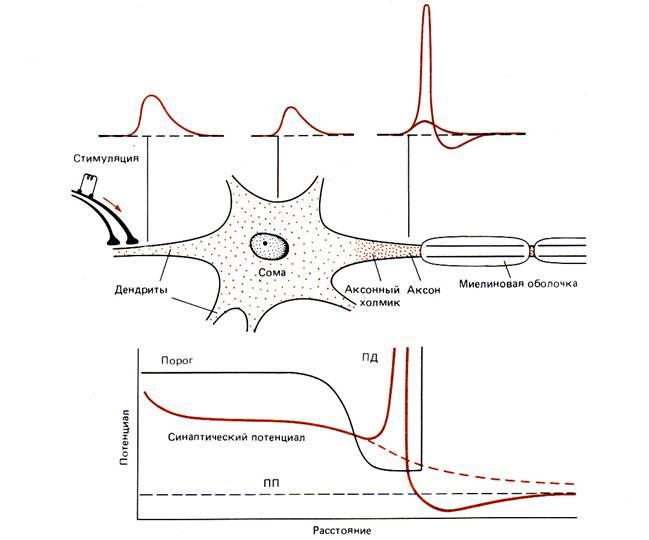
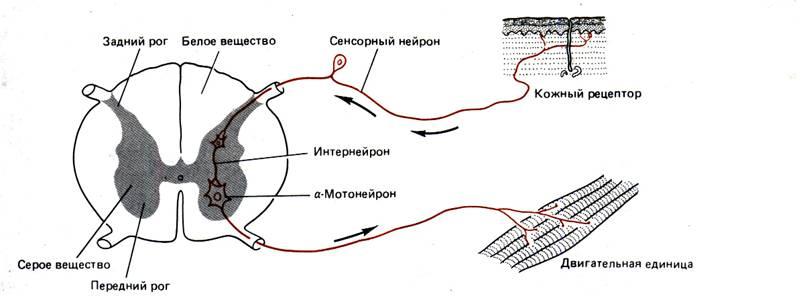
На рис. 6–2 схематично изображен мотонейрон (двигательный нейрон) позвоночных. Тела таких нейронов лежат в спинном мозге, а отростки иннервируют волокна скелетных мышц. Это – классический нейрон типа 1, специализированный для проведения возбуждения на большие расстояния и входящий обычно в состав контуров, объединяющих отдаленные структуры (рис. 6–3). На мембране дендритов (древовидных цитоплазматических выростов мембраны) и сомы (тела) таких нейронов оканчиваются отростки других нервных клеток. По аксону потенциалы действия (ПД) проходят от зоны генерации ПД, расположенной рядом с аксонным холмиком, к окончаниям, которые в нашем примере иннервируют мышечные клетки. Нейронные отростки – дендриты и аксоны – вырастают из сомы в процессе развития, и по ним идет медленный, но постоянный ток белков и других веществ, образующихся в теле клетки. Будучи отделенными от тела, эти отростки постепенно перерождаются и через несколько дней или недель погибают. У млекопитающих регенерация (восстановление) аксонов наблюдается только в периферических нервах; у низших же беспозвоночных регенерация и реиннервация мышц происходят довольно легко.
|
|
|
Рис. 6.2. Мотонейрон спинного мозга позвоночного. Указаны функции, выполняемые различными его частями. Направление передачи сигналов изображено цветными стрелками.
|

|
|
|
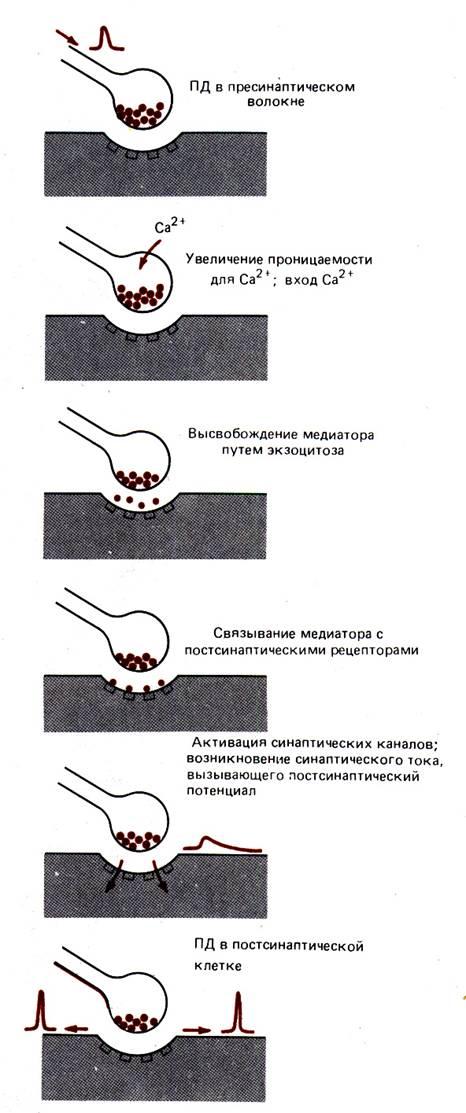
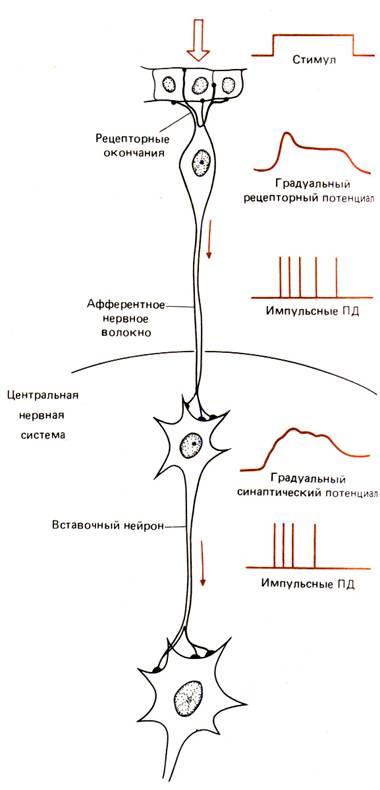
Рис. 6.3. Области возникновения градуальных и импульсных электрических сигналов в нейронной цепи: Градуальные потенциалы, возникающие в чувствительных окончаниях афферентных (чувствительных, сенсорных) нервных клеток в ответ на раздражитель, приблизительно соответствуют его величине и длительности, хотя они и не бывают строго пропорциональным амплитуде раздражителя и не повторяют его конфигурацию. Эти потенциалы распространяются по телу чувствительного нейрона и вызывают в его аксоне импульсные распространяющиеся потенциалы действия. Когда потенциал действия достигает окончания нейрона, происходит выброс медиатора, приводящий к появлению градуального потенциала в следующем нейроне. Если в свою очередь этот потенциал достигает порогового уровня, в этом постсинаптическом нейроне появляется потенциал действия или серия таких потенциалов. Таким образом в нервной цепи наблюдается чередование градуальных и импульсных потенциалов.
|

Функция нейрона зависит от его структуры и от свойств поверхностной мембраны. К этим свойствам в свою очередь относятся пассивные электрические характеристики –емкость и сопротивление (гл. 5), а также наличие различных ионных каналов с воротами, определяющими активное поведение мембраны. Эти ионные каналы распределены по поверхности нейрона неравномерно; они сосредотачиваются в различных его участках, выполняющих соответственно различные специализированные функции. Так, в мембране аксона встречаются преимущественно быстрые потенциалзависимые натриевые каналы, отвечающие за проведение нервных импульсов в большинстве нейронов (в частности, в нейронах типа I). В окончаниях аксона содержатся потенциалзависимые кальциевые каналы и другие специализированные структуры, отвечающие за выделение медиаторов во внеклеточную среду. Такое выделение происходит в области особых соединений синапсов, посредством которых нейроны передают сигналы на свои клетки–мишени. В мембране дендритов и сомы нервных клеток имеются каналы, активируемые теми медиаторами, которые выделяются окончаниями других нейронов. При активации этих каналов возникают постсинаптические токи; эти токи интегрируются (алгебраически суммируются), и в результате возникают постсинаптические потенциалы в дендритах, теле и аксонном холмике нейрона. Таким образом, различные отделы нейрона специализированы как с анатомической, так и функциональной точек зрения.
На долю глиальных клеток, или нейроглии, представленных различными типами клеток, приходится около половины общего объема нервной системы у позвоночных (у большинства беспозвоночных – меньше). Глиальные клетки заполняют все межнейронное пространство, за исключением очень узких (~ 20 нм) промежутков между этими клетками и мембранами нейронов.
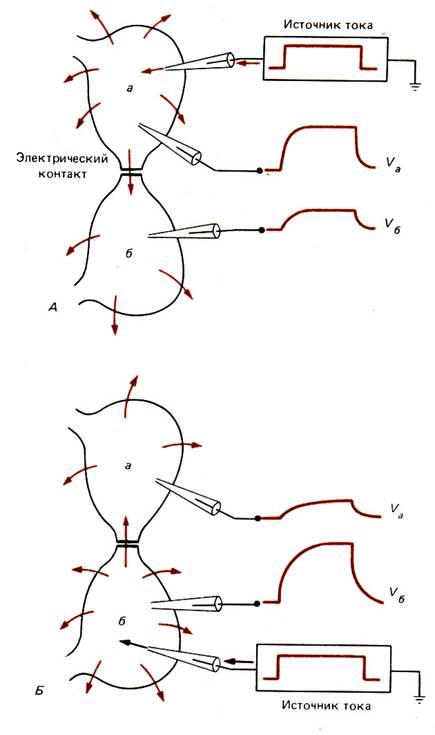
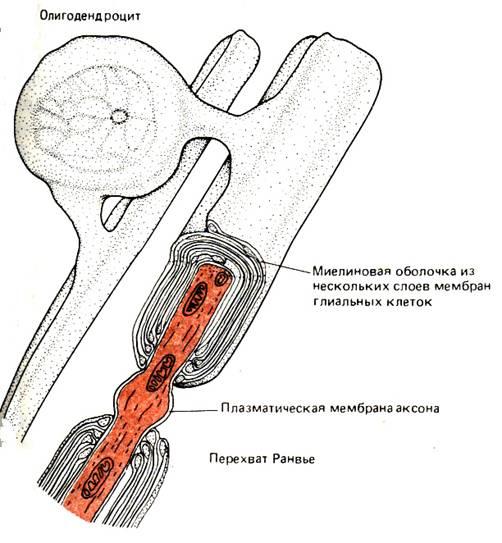
В глиальных клетках иногда встречаются потенциалзависимые ионные каналы, однако обычно они не генерируют ПД. Таким образом, эти клетки электрически невозбудимы, и их роль в деятельности нервной системы долгое время оставалась неясной. Мембраны глиальных клеток высокопроницаемы для К+, и граничащие друг с другом глиальные клетки часто электрически сопряжены, что обеспечивает переход К+ от одной клетки к другой. Благодаря такому транспорту глиальные клетки могут снижать локально высокую концентрацию К+, которая создается в узких межклеточных промежутках, когда этот ион выходит из нейронов при возбуждении. В дальнейшем К+ медленно выделяется из глии и вновь захватывается нейронами. По–видимому, в этом заключается одна из функций глии по обеспечению деятельности нервных клеток. Поскольку потенциал покоя клетки частично зависит от внеклеточной концентрации К+ (см. рис. 5–13), электрическая регистрация потенциалов глиальных клеток оказалась удобным способом, позволяющим следить за изменением внеклеточной концентрации этого иона, происходящим при возбуждении нейронов. Деполяризация мембран глиальных клеток, обусловленная накоплением К+ во внеклеточной среде при возбуждении нейронов, сохраняется в течение нескольких секунд, и это указывает на удаление избыточного К+ из узких внеклеточных промежутков. Очевидно, по крайней мере отчасти калий удаляется через поверхностные мембраны как нервных, так и глиальных клеток. «Забуферирование» резких подъемов содержания К+ во внеклеточной жидкости, осуществляемое многочисленными глиальными клетками, препятствует накоплению этого иона во внеклеточном пространстве. Если бы этого не было, то из–за повышения концентрации К+ во внеклеточной среде нейроны сильно деполяризовывались бы, а это нарушало бы такие функции, как генерация нервного импульса и выделение медиаторов в синапсах. Глиальные клетки не только служат «буферами калия», но также обеспечивают нейронам структурную и, возможно, метаболическую поддержку. Как мы вскоре увидим, глиальные клетки двух типов – шванновские клетки и олигодендроциты – еще и образуют вокруг аксона изолирующую оболочку.