

Первинна будова кореня
Корінь первинної будови ззовні вкритий первинною покривною тканиною ризодермою, або епіблемою. Як і епідерма надземних органів вона виконує декілька функцій, найважливішою з яких є функція поглинання з ґрунту води і розчинів мінеральних солей та передача їх клітинам корової паренхіми. Ця тканина складається з двох типів клітин - тих, що перетворюються на кореневі волоски, і тих, що залишаються незмінними.
Кореневі волоски - це справжні вирости клітин ризодерми, вони не відмежовані від неї перетинками. Завдяки кореневим волоскам поглинальна поверхня кореня збільшується в 6-12 разів, а загальна їхня кількість у рослин може досягати кількох мільярдів. Довжина кореневих волосків, які закінчили ріст, у різних рослин коливається від 0,05 до 10,0 мм. Самі довгі кореневі волоски у цукрового буряка.
Кореневі волоски формують не всі клітини ризодерми. Клітини, які здатні утворювати кореневий волосок, називають трихобластами. Вони коротші і мають густу цитоплазму.
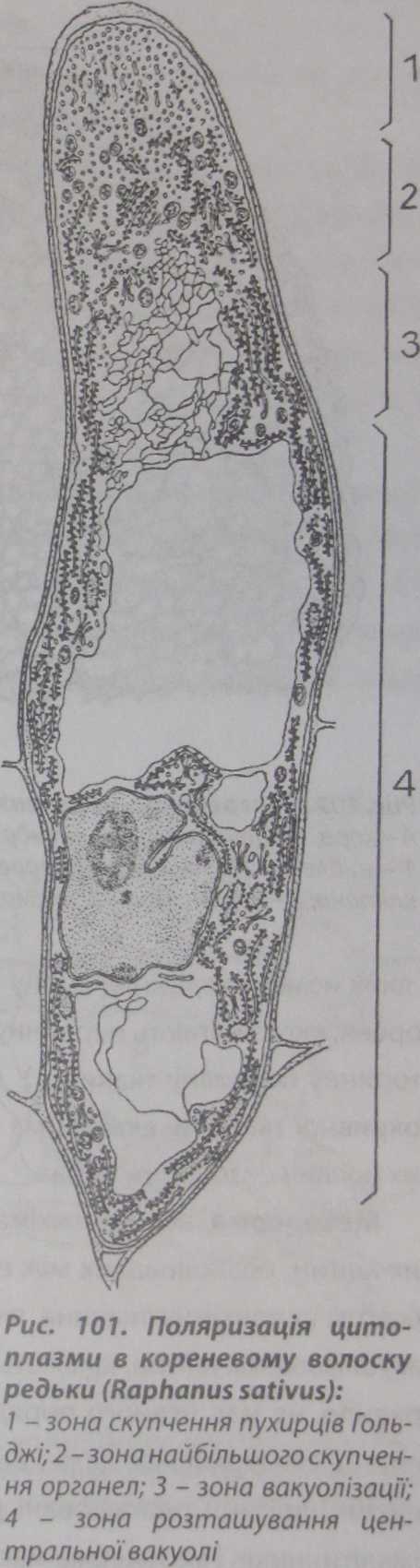
Кореневі волоски мають тонкі целюлозні стінки, а їхня верхівка складається майже з чистого пектину і геміцелюлоз. Біологічне значення такого складу полягає в тому, що пектин не перешкоджає росту волоска. На початку онтогенезу клітини кореневого волоска спостерігається поляризація його цитоплазми, при цьому виділяють чотири зони, залежно від метаболічної активності: 1 - зону скупчення пухирців Гольджі; 2 - зону скупчення органел;
З - зону вакуолізаціїта 41 зону центральної вакуолі (рис. 101).
Кореневі волоски - недовговічні утворення. Тривалість їхньої життєдіяльності не перевищує 10-20 днів. У міру росту кореня на кінчику утворюються нові волоски, а на верхній межі старі кореневі волоски відмирають - у міру віддалення від апекса вони вкорочуються, втрачають функціональну активність, відмирають і злущуються.
Кореневі волоски здатні виділяти особливі кислоти, за допомогою яких вони розчиняють ґрунтові частинки і міцно приклеюються до них, тим самим сприяючи закріпленню кореня у ґрунті.
 Центральна
частина кореня складається з первинної
кори і осьового циліндра.
Центральна
частина кореня складається з первинної
кори і осьового циліндра.
Первинна кора кореня складається з типовоїпаренхіми. В ній помітно три шари: зовнішній - екзодерму, середній - мезодерму і внутрішній - ендодерму (рис. 102).
Клітини екзодерми розташовані безпосередньо під ризодермою. У різних рослин екзодерма буває одношаровою, двошаровою і багатошаровою. Клітини екзодерми живі, щільно зімкнені, багатокутні, їхні стінки сильно потовщені, вони більші за розмірами і витягнуті по радіусу, чим відрізняються від розташованих глибше клітин.
Екзодерма замінює ризодерму у тій частині кореня, яка втрачає функцію поглинання і де утворюються бічні корені. Морфологічно ця частина кореня займає проміжне положення між первинною і вторинною будовами кореня. Як правило, після відмиранням клітин ризодерми клітини екзодерми корковіють. У кільці окорковілих клітин екзодерми звичайно залишаються поодинокі клітини з целюлозними стінками. Ймовірно, вони забезпечують зв'язок із зовнішнім середовищем.
Слід зазначити, що екзодерма особливо характерна для однодольних рослин,
Рис.
102. Поперечний зріз кореня Ірису (Ігі$):
А
- кора,
В
- центральний
циліндр:
1
- епіблема;
2 - поглинальна паренхіма (мезодерма);
3
- ендодерма;
4
- пропускна
клітина; 5
- пери
цикл; б
- ксипема;
7 - флоема; 8
- екзодерма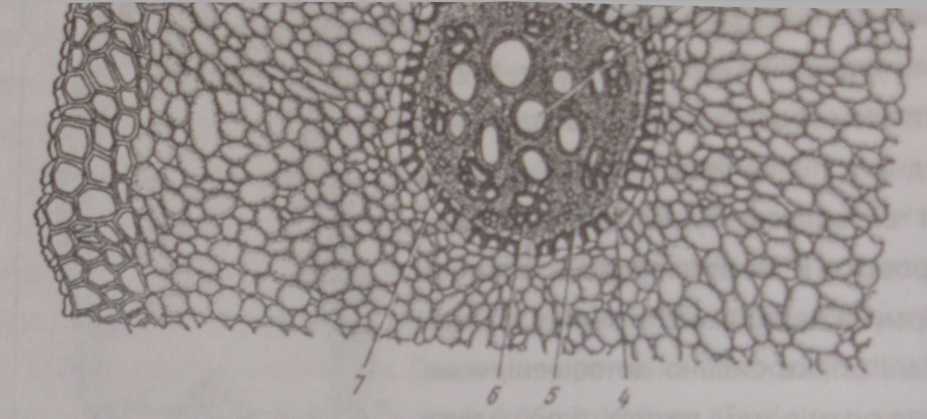
у яких немає вторинних змін у корені. У цьому випадку для старих частин кореня, які зберігають первинну будову, окорковіла екзодерма ніби заміняс вторинну покривну тканину. У дводольних рослин, у разі появи вторинної покривної тканини, екзодерма відмирає, розривається і злущується. У водних рослин екзодерми немає.
Мезодерма, або паренхіма первинної кори, - це сукупність паренхім- них клітин, розташованих між екзодермою і ендодермою. Це найбільша за обсягом паренхімна тканина, яка складається майже з однакових округлих вакуолізованих клітин, кількість шарів яких варіює. Розташування клгтин, як правило, не має певного порядку. Нещільне розташування основно? маси клітин мезодерми має важливе знамення для аерації. У водних рослин па* ренхімні клітини розташовані радіально, що зумовлює розвиток великих міжклітинників І мезодерма має вигляд аеренхіми У деяких рослин повітря*
мі порожнини розташовані безперервно від кореня до листка, що забезпечує надходження кисню до підземних органів.
Мезодерма транспорту* водний розчин мінеральних солей до цен- іраяьиого циліндра кореня у горизонтальному напрямку.
Ендодерма
~ внутрішній шар первинної кори кореня,
вона складається
і одного ряду клгтин, який кільцем оточує центральний циліндр. Дуже рідко ендодерма бувас двошаровою, наприклад у конвалії і лимонника. Ендодермою регулюється надходження води у горизонтальному напрямку від периферії до центрального циліндра, де розташовані провідні елементи. Біль* «юь клітин ендодерми мас потовщені, практично непроникні здерев'янілі «яітмині стінки - пояски Каспар* (рис 103).
З
наявністю поясків
Каспарі повя іують активну участь
ендодерми
в
регулювали радіального
транспортування речовин тканинами
кореня. Пояс- т
Катарі, завдяки
гідрофобним властивостям речовин, що
входять до їхнього
складу* перекривають
на рівні ендодерми вільний простір
клітинних сіікж
і перешкоджають
дифузному пересуванню речовин по
стінці, Внасл*
док з'єднання ппазмалеми з клітинною стінкою перекривається також і пе- риплазматичний простір. Таким чином, пояски Каспарі стають ефективним бар'єром на шляху пасивного пересування речовин апопластом. Припинення вільної дифузії речовин по апопласту сприяє їхньому активному пересуванню симпластним шляхом клітинами ендодерми і корової паренхіми.
Із переходом кореня до вторинної будови, ендодерма також переходить до наступної стадії розвитку, яка супроводжується відкладенням на первинну стінку суберину.
Суберинізація здійснюється, як правило, одночасно у більшості клітин ендодерми, розташованих на одному рівні кореня, але деяка кількість клітин запізнюється. У зв'язку з цим, серед клітин ендодерми із суберенізованими стінками зустрічаються клітини, які зберігають початковий стан стінок - пропускні клітини. Вони забезпечують здійснення контрольованого радіального транспорту речовин від кореневих волосків і паренхіми до центрального циліндра кореня. Всі пропускні клітини розташовані проти елементів ксиле- ми центрального циліндра.
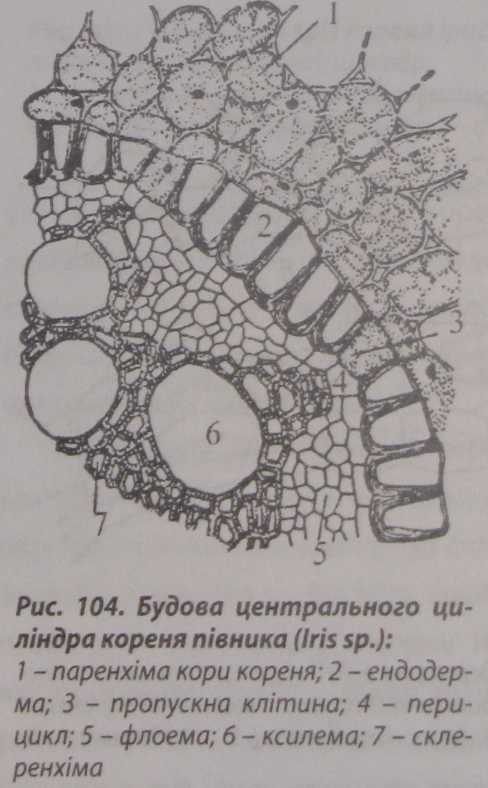 Центральний
циліндр кореня
з первинною будовою в основному
складається з судин і ситоподібних
трубок. Він відмежований від первинної
кори одношаровою латеральною меристемою
- перициклом (рис. 104).
Центральний
циліндр кореня
з первинною будовою в основному
складається з судин і ситоподібних
трубок. Він відмежований від первинної
кори одношаровою латеральною меристемою
- перициклом (рис. 104).
Зовнішні епідермальні залозки: трихоми, волоски, сольові залозки. 161
Первинна будова стебла 174
Вторинна будова стебла дводольних трав'янистих рослин 55
Будова стебла водних рослин 190
АНАТОМІЧНА БУДОВА ЛИСТКА 191
Анатомічні особливості різних типів коренів 230
АНАТОМІЧНА БУДОВА НАСІНИНИ 112
АНАТОМІЧНА БУДОВА ПЛОДІВ 122
редукції являє собою переривчастий перицикл, характерний для злакових, коли клітини перициклу напроти ксилемних полюсів диференціюються як трахеальні елементи, які безпосередньо межують з внутрішнім шаром первинної кори - ендодермою.
Завдяки діяльності перициклу утворюються бічні корені та адвентивні бруньки. В деяких випадках перицикл виконує запасаючу функцію (у складноцвітих в клітинах перициклу накопичується молочний сік, а у моркви - олії), а то і механічну (у деяких однодольних клітини перициклу дерев'яніють).
Самий центр кореня складається з провідних елементів. Флоема та кси- лема тут чергуються між собою і розташовані радіальними групами - радіальний провідний пучок. Його утворенню передує закладання прокамбію у вигляді центрального тяжа. Диференціація клітин прокамбію в елементи протофлоеми, а потім і протоксилеми починається на периферії, у подальшому ці тканини розвиваються доцентрово.
Перші провідні елементи ксилеми в корені формуються з прокамбію ек- зархно, тобто першочергово сформовані судини розташовані на зовнішньому боці ксилеми, а наймолодші - ближче до центра.
У стеблі, на відміну від кореня, трахеальні елементи формуються ен- дархно.
Первинна флоема кореня розвивається, як і ксилема, екзархно: самі ранні елементи флоеми знаходяться біля перициклу, а молодші - ближче до центра.
Залежно від кількості променів флоеми та ксилеми розрізняють декілька типів коренів: монархні - однопроменеві, диархні - двопроменеві, три-, тетра-, пента-, текса-, поліархні. Кількість променів флоеми та ксилеми у деяких рослин є порівняно сталою величиною. У більшості рослин цей показник може змінюватися залежно від екологічних умов, особливо за дії екстремальних факторів.
Механічна частина кореня первинної будови представлена склеренхімними волокнами, які майже завжди розташовані в центрі осьового циліндра, в той час як у стеблі такі тканини розташовані на периферії. Таке розташування залежить від різних функцій цих органів.
Між коренем і стеблом розташована зона, яку називають кореневою шийкою. Це перехідна зона. У рослин з первинною будовою у кореневій
шийці ксилема поступово переміщується з колатерального положення стеблових пучків у радіальне, властиве кореням. Кількість ксилемних пучків кореня відповідає кількості судинних пучків стебла. Після переходу кореня до вторинної будови його структура стає подібною до вторинної будови стебла, тому у кореневій шийці відбувається пряме з'єднання обох осьових органів.
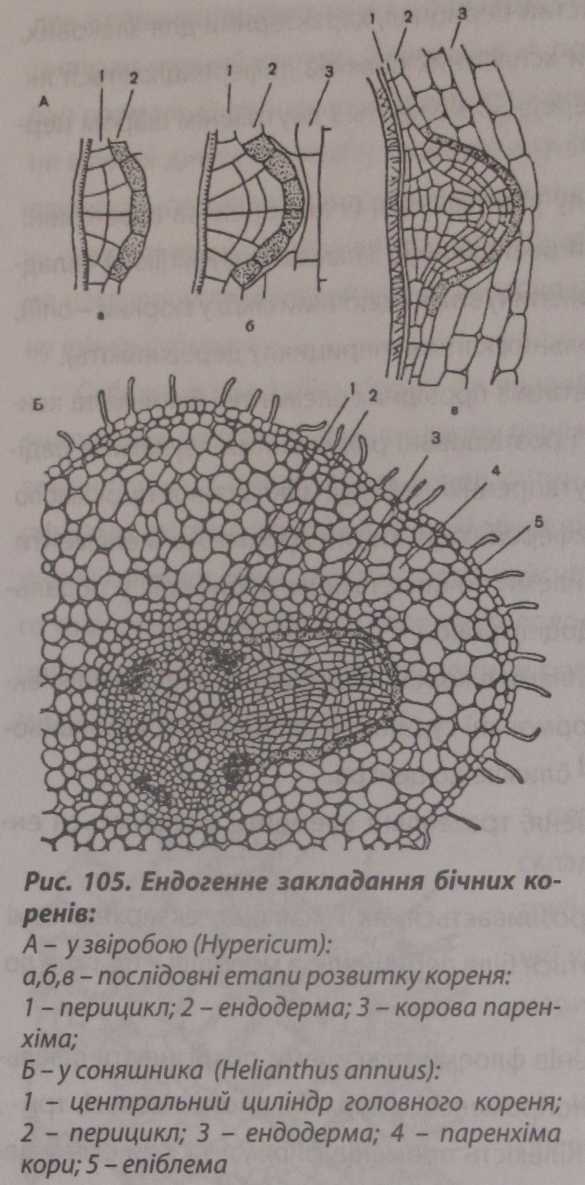 Закладання
бічних коренів. Бічні
корені закладаються в спеціалізованій
меристематич- ній тканині - перициклі,
найчастіше проти груп ксилемних
елементів осьового циліндра (рис. 105). У
злаків бічні корені закладаються проти
флоемних ділянок. Спочатку кілька клітин
перициклу видовжуються в радіальному
напрямку, а через певний час поділ
відбувається тангентально, завдяки
чому утворюється коренерідна
дуга, яка
є основою апікальної меристеми бічного
кореня. Клітини цієї дуги діляться.
Закладання
бічних коренів. Бічні
корені закладаються в спеціалізованій
меристематич- ній тканині - перициклі,
найчастіше проти груп ксилемних
елементів осьового циліндра (рис. 105). У
злаків бічні корені закладаються проти
флоемних ділянок. Спочатку кілька клітин
перициклу видовжуються в радіальному
напрямку, а через певний час поділ
відбувається тангентально, завдяки
чому утворюється коренерідна
дуга, яка
є основою апікальної меристеми бічного
кореня. Клітини цієї дуги діляться.
Одночасно з формуванням коренерідної дуги починається поділ клітин ендодерми, які розташовані ззовні від дуги. Завдяки цьому поділу ендодер- мальний поясок розширюється і не заважає росту бічного кореня. З цієї частини ендодерми формується коренева кишенька (чохлик). Розвиваючись у довжину, бічний корінь прокладає шлях через первинну кору та ризодерму за допомогою кишеньки, яка розсуває клітини паренхіми кори, або виділяє
ферменти, які руйнують клітини кореня. Коли бічний корінь виходить назовні, кишенька зникає, а апекс бічного кореня формує новий чохлик.
Бічні корені рослин утворюються у певному співвідношенні до потужності провідних тканин материнського кореня. Найчастіше вони утворюються у внутрішніх тканинах підземного органа і на відносно молодих його ділянках, що має пристосувальне значення. Якби галуження відбувалося в апексі кореня, то це ускладнювало б його просування в ґрунті.
Вторинна будова кореня. Первинна будова кореня характерна для однодольних та деяких дводольних рослин.
У більшості дводольних властива молодим рослинам первинна будова кореня згодом набуває вторинної будови.
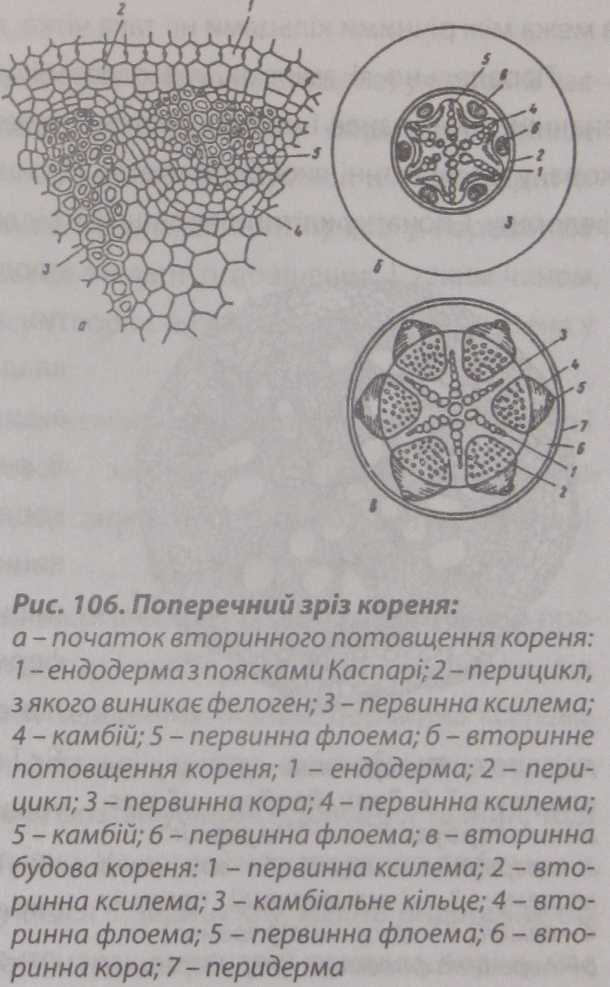 Вторинні
зміни у корені починаються вище зони
бічних коренів, з моменту закладання
камбіального кільця. Камбій формується
з паренхімних клітин, розташованих між
флоемою та ксилемою. В результаті між
групами первинної флоеми та ксилеми
виникає дугоподібна смужка камбію, яка
огинає кси- лему з зовнішнього боку, а
флоему-з внутрішнього (рис. 106).
Вторинні
зміни у корені починаються вище зони
бічних коренів, з моменту закладання
камбіального кільця. Камбій формується
з паренхімних клітин, розташованих між
флоемою та ксилемою. В результаті між
групами первинної флоеми та ксилеми
виникає дугоподібна смужка камбію, яка
огинає кси- лему з зовнішнього боку, а
флоему-з внутрішнього (рис. 106).
Окремі смужки в результаті антиклінального поділу клітин подовжуються і зливаються, утворюючи суцільну смужку камбію, яка на поперечному зрізі має зірчасту форму.
Подальша діяльність утвореного камбію в корені така сама, як і в стеблі: до центра він відкладає вторинну ксилему, а до периферії | флоему. Разом
з тим, з камбію утворюються паренхімні клітини серцевинних променів, які розташовані
у вигляді прошарку між флоемою і ксилемою. Діяльність камбію інтенсивніше в тій частині, де формується ксилема. Внаслідок цього, проміжки між радіальними рядами ксилеми і флоеми вирівнюються, і камбій на поперечному зрізі набуває вигляду кільця.
Вирівнювання кільця камбію, утворення ним концентричних шарів лубу і деревини і вторинних серцевинних променів зумовлює подібність вторин- ноїструктури кореня з будовою стебла. Основна причина цієї подібності полягає в тому, що ця частина кореня виконує ту ж функцію перенесення води і органічних речовин, що і стебло.
Після формування вторинних тканин первинна флоема кореня зміщується до периферії, а первинна ксилема - до центра.
Діяльність камбію в коренях характеризується періодичністю. Розрізняють два періоди: весняний (березень-червень) і осінній (з вересня до початку листопада). Річні прирости деревини в корені значно вужчі, ніж у стеблі, а межа між річними кільцями не така чітка, як у стеблі.
Паралельно зі змінами, які спостерігаються у центральному циліндрі, значних змін зазнає і корова частина кореня. З'являється нова - вторинна кора, у формуванні якої бере участь перицикл. Перицикл виконує функцію фелогену. Спочатку клітини перициклу відділяють назовні клітини, які також
 продовжують
ділитись. Цей шар клітин виконує
функцію фелогену і відкладає до
периферії клітини корка, а до середини
- клітини фелодерми.
продовжують
ділитись. Цей шар клітин виконує
функцію фелогену і відкладає до
периферії клітини корка, а до середини
- клітини фелодерми.
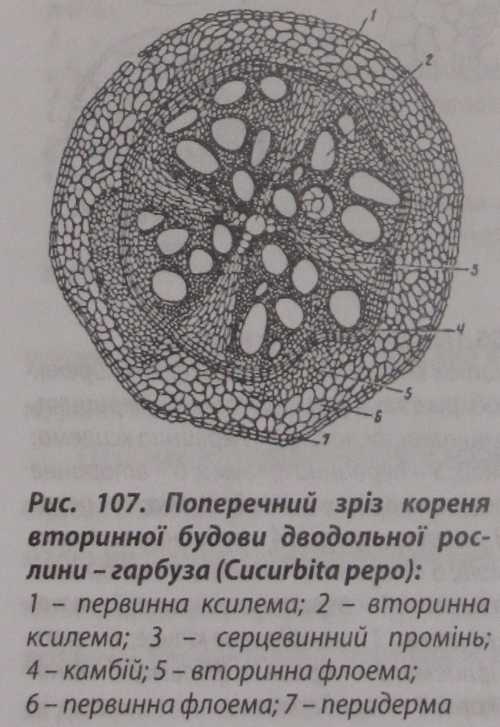
В результаті формується перидерма кореня, яка утворюється з клітин первинної латеральної меристеми - перициклу, на відміну від стебла, де вона формується з шару клітин, що лежать під епідермою, або з самої епідерми.
Як і в стеблі, вона розростається і потовщується. При цьому зовнішні від неї тканини первинної кори відмежовуються, відмирають, швидко руйнуються мікрофлорою ґрунту (рис. 107).
Перидерма кореня, так само як і перидерма стебла, має сочевички такої ж будови, але значно більшого розміру.
Товщина шару вторинної кори кореня менша, ніж стебла. Вторинна кора кореня складається з луб'яної паренхіми, в якій часто накопичуються запасні поживні речовини. До корової паренхіми прилягає флоема.
Отже, характерні етапи переходу кореня від первинної до вторинної будови такі: поява камбію між ділянками первинної флоеми і ксилеми, утворення фелогену із перициклу з наступним формуванням вторинної кори, злущування первинної кори, заміна радіального розташування провідних елементів на колатеральне.
Вторинні зміни в корені однодольних зустрічаються рідко. У трав'янистих однодольних первинна будова кореня зберігається до закінчення життя. З часом клітини ендодерми, більшість клітин мезодерми і паренхіми центрального циліндра склерифікуються (за рахунок лігніфікації і мінералізацій.
У багатьох деревних однодольних (пальми, юка, драцена) у коровій частині кореня із паренхімних або перициклічних клітин формується кільце твірної тканини - кільце потовщення, з якого формується кільце закритих судинних пучків. За сформованим кільцем судинних пучків у коровій паренхімі, на її периферії формується нове кільце потовщення. І, таким чином, в результаті багаторазового процесу утворюється вторинна будова кореня у деревних однодольних.
У тих багаторічних рослин, у яких немає кілець потовщень, ендодерма і перицикл дерев'яніють або корковіють і заміняють вторинну покривну тканину кореня. У цьому разі в осьовому циліндрі вторинних змін не спостерігають.
Отже, корінь і стебло - це осьові органи, які мають радіальну або променеву симетрію, верхівковий тип росту і здатні до галуження. Корінь від стебла відрізняється розташуванням первинних тканин: первинна ксилема в ньому екзархна, тобто розташована далі від центра, а в стеблі вона ендарх- на. Для кореня характерне радіальне розташування провідних тканин, тоді як у стеблі вони сконцентровані у пучки. На корені не утворюються бруньки, листки і квітки. Бічні корені формуються з перициклу, мають ендогенне походження і закладаються в глибоких шарах кореневої тканини. Корінь має
кореневий чохлик, якого немає в стеблі, має кореневі волоски, які вбирають воду і мінеральні сполуки. Корінь завжди має ендодерму, якої в стеблі часто немає.
Корінь вторинної будови немає первинної кори.