

Вторинна будова стебла дводольних трав'янистих рослин
Основною особливістю будови стебла дводольних трав'янистих рослин є посилений розвиток паренхімних клітин, які переважають у серцевині, корі і серцевинних променях. Крім того, порядок розташування механічних тканин у цих рослин теж специфічний. Як правило, під покривною тканиною в поверхневих шарах кори закладається механічна тканина, а саме коленхіма.
Вторинна будова стебла дводольної трав'янистої рослини мало відрізняється від його первинної будови. Вторинні зміни у стеблі з'являються з початком функціонування камбіального кільця, завдяки чому потовщується стебло.
Відомо, що розрізняють два типи вторинної будови осьових органів: пучковий і безпучковий.
Прикладом пучкового типу вторинної будови стебла трав'янистих рослин є конюшина повзуча (ТгіїоНит герепз) (рис. 78). Стебло вкрите епідермою, стінки клітин якої потовщені і кутинізовані. Під епідермою розташована корова тонкостінна паренхіма. Паренхімні клітини первинної кори містять хлоропласти, здатні до фотосинтезу. Згодом клітини корової паренхіми, які розташовані безпосередньо під епідермою, потовщуються і перетворюються на коленхіму, яка надає стеблам трав'янистих рослин еластичності і пружності.
Осьовий циліндр розвинутий значно сильніше, ніж корова частина. Це характерно для більшості дводольних. Ендодерма не виражена, не видно і диференційованого перициклу.

Основні елементи осьового циліндра - судинно-волокнисті пучки і добре розвинена крупноклітинна серцевинна паренхіма. Пучковий камбій добре виражений, міжпучкового поки немає, він утворюється дещо пізніше з клітин серцевинних променів. В результаті формується суцільне камбіальне кільце. Пучковий камбій утворює вторинні елементи флоеми і ксилеми, а міжпучковий камбій - формує серцевинні промені. Тому окремі судинно- волокнисті пучки не можуть зблизитись і залишаються розмежованими протягом усього життя рослини.
Прикладом стебла з непучковою будовою може бути льон вузьколистий (іИпит и$іШІ5$ітит) (рис. 79). У корі і центральній частині стебла з непучковою будовою важко встановити межу між первинними та вторинними елементами. Клітини первинної паренхіми кори залишаються майже незмінними - відносно дрібного розміру і видовженої форми внаслідок розтягування
стебла на ранніх етапах розвитку рослини у горизонтальному напрямку. Добре виражені ділянки товстостінних луб'яних волокон камбіального походження, під якими суцільною зоною розташовані елементи вторинної флоеми, утворені кільцем камбію.
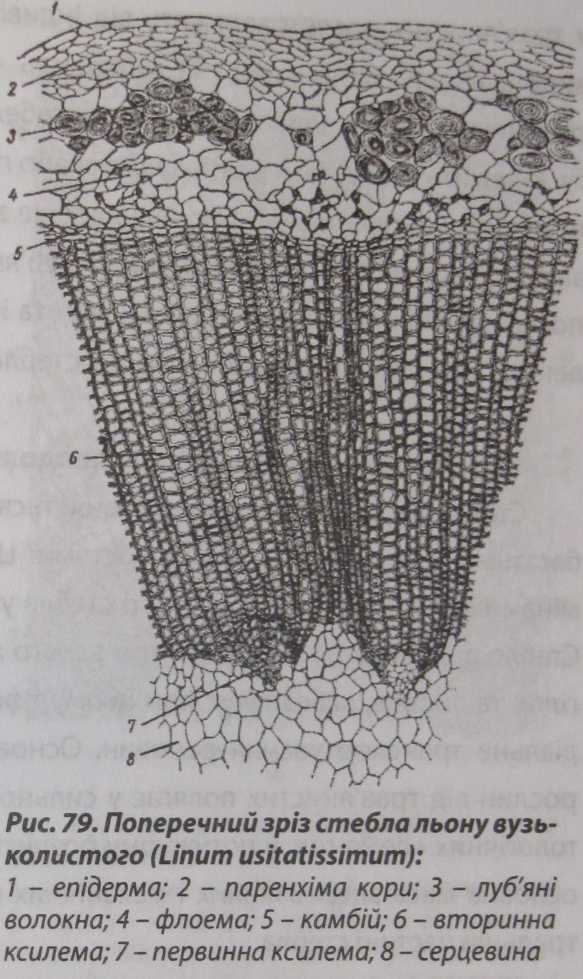 Ксилемна
зона вторинного походження відносно
широка та однорідна. В її основі можна
помітити ділянки первинної ксилеми, що
частково заходять у серцевину. Нові
елементи ксилеми і флоеми відкладаються
суцільним кільцем: флоема до периферії,
а ксилема до середини. Подекуди камбій
відкладає з обох боків паренхімні
клітини серцевинних променів.
Ксилемна
зона вторинного походження відносно
широка та однорідна. В її основі можна
помітити ділянки первинної ксилеми, що
частково заходять у серцевину. Нові
елементи ксилеми і флоеми відкладаються
суцільним кільцем: флоема до периферії,
а ксилема до середини. Подекуди камбій
відкладає з обох боків паренхімні
клітини серцевинних променів.
У процесі поділу камбіальних клітин відбувається потовщення стебла, його розростання в радіальному і тангентальному напрямках. При цьому всі первинні анатомічні структури, розташовані глибше від камбію, перебувають у стеблі в такому ж стані, в якому вони були до початку вторинного росту. А ті первинні тканини, що лежать назовні від камбію (флоема, перицикл, ендодерма, кора, епідерма), завдяки його діяльності відсуваються вторинними тканинами до периферії органа, після чого збільшення товщини стебла перевищує пластичні можливості цих тканин і вони, як правило, сплющуються з неминучими радіальними розривами. На початку вторинного росту розривається і перестає функціонувати ендодерма. Значно довше протидіють розривам перицикл і кора, бо вони мають тонкостінні, здатні до поділу клітини. Отже, вторинні зміни у стеблі дводольних з'являються з початком фор-
у трав'янистих рослин залежить від індивідуальних властивостей та умов вирощування. Наприклад, на швидкість формування вторинної будови рослини суттєво впливає і здатність стебел до галуження, що, в свою чергу, залежить від густоти розташування або посіву. Збільшення густоти посіву луб'яних культур (льону, конопель) довше зберігає первинну будову стебел, за якої утворюються цінніші первинні луб'яні волокна. Навпаки, в зріджених посівах бавовнику, рицини, соняшнику та ін. збільшується галуження, посилено утворюються вторинні тканини, стебла потовщуються і дерев'яніють.
Вторинна будова стебла дводольних деревних рослин Стебло деревних рослин відрізняється від стебла трав'янистих рослин багатьма структурними особливостями. Це визначається насамперед відмінами функціональної діяльності стебла у трав'янистих і деревних рослин. Стебло деревних рослин протягом всього життя несе на собі великий тягар гілок та листків, забезпечує при цьому ефективне висхідне, низхідне та радіальне транспортування речовин. Основна відмінність стебла деревних рослин від трав'янистих полягає у сильному здерев'янінні (лігніфікаці'О гістологічних елементів, у потужному розвитку механічних тканин, при цьому основна маса здерев'янілих і механічних елементів зосереджується в центральній частині стебла.
У стеблах дводольних рослин вторинні зміни настають дуже рано. На відміну від трав'янистих, у деревних дводольних вторинні структури з'являються дуже швидко і елементи первинної будови ніби розчиняються у масі вторинних утворень. У1-2-3 річних гілок поверх перидерми можна спостерігати залишки відмерлої епідерми, у корі - коленхіму, іноді ендодерму. Ці структурні елементи первинної будови згодом перемішуються з вторинними елементами стебла.
У більшості дводольних деревних рослин стебла мають безпучкову будову, тому в апікальній меристемі стебла закладається суцільне прокамбіальне кільце, яке утворює кільце флоеми і кільце ксилеми. Між ними зберігається функціонуючий прошарок прокамбію, який дає початок камбіальному кільцю.
Стебло деревних рослин має таку топографію: первинна кора, до складу якої відносять і покривну тканину, і центральний циліндр, який складається
З самого початку формування вторинної будови стебла деревних рослин зовнішнім шаром кори є перидерма.
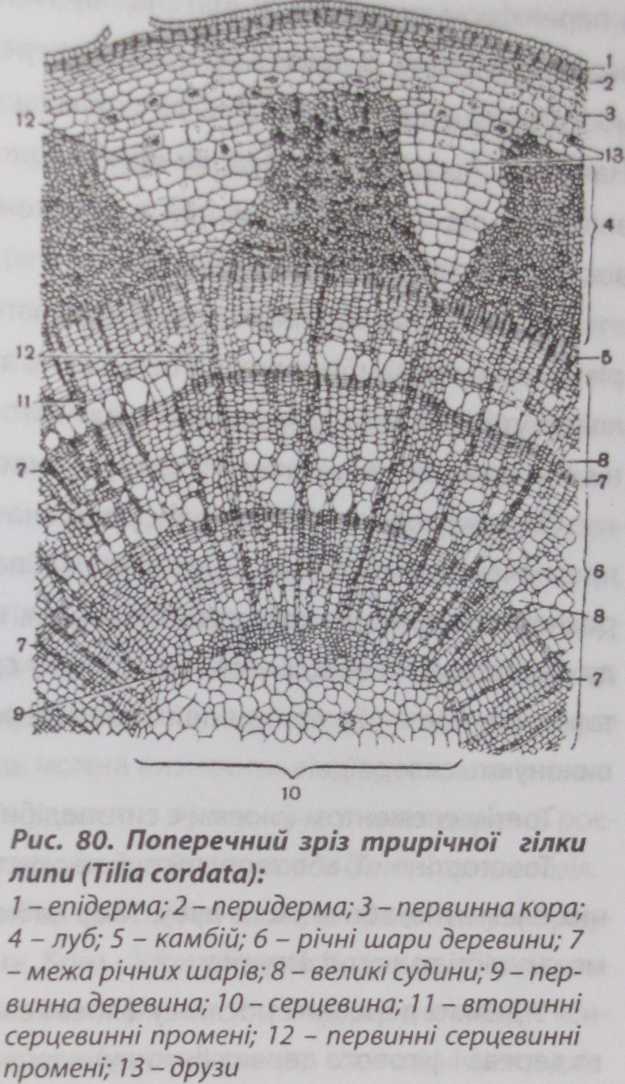 У
багатьох деревних порід перидерма
з часом замінюється, перетворюється на
кірку. Під перидермою до осьового
циліндра розташована корова паренхіма.
Внутрішню межу кори формує первинна
дрібноклітинна паренхіма.
У
багатьох деревних порід перидерма
з часом замінюється, перетворюється на
кірку. Під перидермою до осьового
циліндра розташована корова паренхіма.
Внутрішню межу кори формує первинна
дрібноклітинна паренхіма.
Корова паренхіма у деревних рослин виконує асиміляційну (зовнішні шари містять хлоропласти) і запасаючу (внутрішні шари) функції. Іноді стінки клітин зовнішніх шарів корової паренхіми потовщуються, і ці клітини перетворюються на коленхіму, яка функціонує лише у перші роки життя гілки. 3 віком і у міру потовщення перидерми коленхімні клітини втрачають механічне значення і, змішуючись з наростаючою масою корової паренхіми, стають непомітними. У випадку відшарування перидерми і виникнення нових кілець фелогену коленхіма скидається разом зі старою перидермою.
Під коровою паренхімою розпочинається вторинна кора - зона флоем- них елементів (лубу). У флоемі деревних рослин є три типи клітин: луб'яна паренхіма, товстостінний луб (механічна система) і ситоподібні трубки з клітинами-сулутниками (провідна система).
Основну масу лубу становить луб'яна паренхіма, яка утворюється внаслідок поділу камбіальних клітин. Луб'яна паренхіма дерев і кущів складається з живих клітин, стінки яких тонші і не дерев'яніють, якщо порівнювати
![]()
з паренхімою деревини. У клітинах луо яної паренхіми накопичуються резервні речовини. У луб'яній паренхімі деревних порід є ділянки кристало- носної паренхіми, у клітинах якої відкладаються кристали оксалату кальцію, Наявність і особливості будови кристалоносної паренхіми мають важливе значення під час таксономічних дослідженнь, при визначенні рівня морозостійкості пагонів багаторічних рослин.
Деякі клітини луб'яної паренхіми мають ніжні й тонкі стінки і мало відрізняються від камбіальних клітин, з яких вони утворилися. Сукупність цих клітин утворює камбіформ, у клітинах якого відкладаються резервні речовини. У вторинній флоемі камбіформ зустрічається рідко.
Система товстостінного лубу утворена двома елементами механічного призначення: луб'яними волокнами та перегородчастим лубом. Перего- родчастий луб мало поширений елемент, він, зокрема, відомий у винограду. Характерною особливістю цього лубу є дуже вузенькі порожнини. Крім товстостінного лубу у вторинній флоемі деревних рослин механічну функцію виконують склереїди.
Третім елементом флоеми є ситоподібні трубки і клітини-супутники.
Товстостінний, або твердий луб чергується з прошарками живих провідних елементів, або м'якого лубу. Тому на поперечному зрізі пагона ми бачимо смугасті трикутні фігури у корі.
У деяких деревних рослин у флоемі є молочники, наприклад каучукового дерева і фігового дерева (інжиру).
Луб відмежовується від деревини прошарком камбію. В результаті діяльності камбію формуються вторинна флоема (екзархно, тобто від центра до периферії) і вторинна ксилема (ендархно). Функціональна активність камбію має сезонний характер. Найактивніший камбій навесні.
Під камбієм розташована вторинна ксилема, або деревина. У деревині розрізняють три групи елементів: паренхімні, механічні і провідні.
Клітини паренхіми деревини живі і мають потовщені стінки. Розташовані рівномірно, відносно правильними радіальними рядами. Виконують запасаючу функцію.
Механічна складова деревини представлена волокнами склеренхіми, а саме лібриформом. Лібриформ утворює основну масу деревини. У деяких багаторічних рослин (винограду, плюща) зустрічається перегородчастий лі-
Ьриформ, волокна якого після закінчення росту в довжину та потовщення клітинних стінок діляться поперечними перегородками, які залишаються тонкими і не дерев'яніють. Взимку у клітинах перегородчастого лібриформу є багато крохмалю, тому він виконує механічну й запасаючу функції.
Провідні елементи деревини - судини, або трахеї.
Усі гістологічні елементи деревини утворюються в результаті диференціації клітин камбію, які активні тільки протягом вегетаційного періоду: його діяльність розпочинається навесні і закінчується восени. Деревину, що виникає навесні, називають весняною, і вона істотно відрізняється від деревини, що утворюється в другій половині вегетації - осінньої. Весняна деревина містить більше провідних елементів, паренхімні елементи крупніші, розташовані пухкіше і мають тонкі стінки. Клітини осінньої деревини, навпаки, дрібніші, мають товсті клітинні стінки; в ній переважають волокна лібриформу, які утворюють темнішу зону річного кільця. Осінні елементи здійснюють в основному механічну функцію, а весняні проводять воду та мінеральні речовини. За кількістю річних кілець можна визначити вік дерева, а за їхньою структурою можна скласти певне уявлення про умови, в яких деревні рослини розвивалися. Структуру річних кілець вивчає наука дендрохронологія.
Періодичність діяльності камбію практично не проявляється у формуванні елементів вторинної флоеми. Крім цієї особливості, для камбію характерна і чітко виражена нерівнозначність кількості утворюваних клітин ксилеми і флоеми: на 100 елементів деревини протягом літа відкладається лише 10 елементів вторинної флоеми.
Найстаріші річні кільця розташовані у центрі стебла. З віком клітини паренхіми деревини починають відмирати, а судинні елементи втрачають провідність. Навіть неозброєним оком на поперечному зрізі стовбура дерева можна помітити дві частини: центральну - темнішого кольору, яку називають ядровою деревиною, і периферійну - молоду, світлішого кольору, яку називають заболонню. У липи або ялини центральна зона деревини майже не відрізняється забарвленням від периферійної. Вона містить живу паренхіму. У цьому випадку центральну частину деревини, що втратила провідну функцію і не має забарвлення, називають спілою деревиною.
Через деревину і вторинну флоему проходять серцевинні промені. Із міжпучкового камбію утворюються елементи первинних серцевинних про-
менів, а завдяки діяльності камбію утворюються вторинні серцевинні промені. Вторинні серцевинні промені відрізняються від первинних ТИМ, що вони не доходять до серцеви- ни, в них відкладається багато крохмалю, особливо восени.
 У
центрі стебла знаходиться серцевина,
по периферії якої розташована перимеду-
лярна зона.
У
центрі стебла знаходиться серцевина,
по периферії якої розташована перимеду-
лярна зона.
Неповторний за красою і структурою анатомічний малюнок деревини маловиразної і не дуже примітної берези карельської (Веіиіа репсіиіа і сагеїіса), або мармурового дерева, яка зрідка трапляється на кам'янистих ґрунтах у суворих водних та температурних умовах Скандинавії, Фінляндії, Карелії, Білорусі. Царська береза, так ще називали цю рослину, бо вироби з її неповторної деревини могли дозволити собі лише царі та інші перші особи держави.
Унікальна візерунчаста текстура деревини берези карельської утворюється в результаті діяльності камбію, здебільшого навпроти серцевинних променів. Він ніби виходить з під контролю й творить клітини деревини, як йому заманеться: не рівненькими концентричними шарами, як у більшості деревних рослин, а спонтанними ділянками більш швидкого і частіше нерівномірного звивистого нарощування деревини на одному боці
стовбура. Як дослідили фізіологи та анатоми рослин, усі аномалії внутрішньої будови деревини берези карельської зумовлені порушенням балансу і кількісного співвідношення фітогормонів як в органах, де вони утворюються, такі в тканинах, куди вони транспортуються, зокрема, в камбіальні зони.
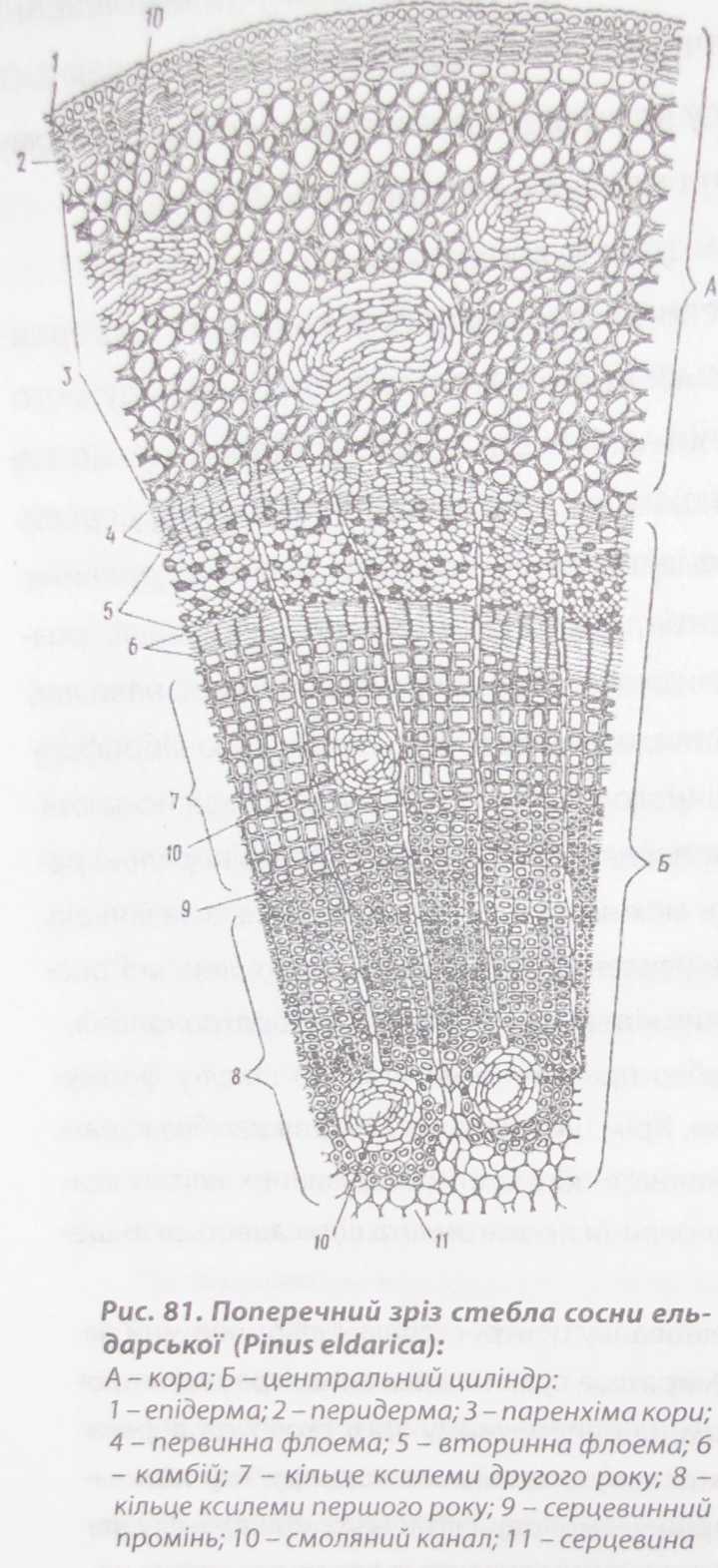
Будова стебла хвойних Анатомічна будова стебла хвойних (голонасінних) рослин має ряд особливостей (рис. 81).
 За
вторинної будови стебло хвойних вкрите
перидермою, яка дуже швидко (4-5 років)
замінюється кіркою. У паренхімі первинної
кори є смоляні ходи, у флоемі немає
клітин-супутників, біля ситоподібних
клітин зустрічаються білкові клітини,
в яких міститься значна кількість
запасних білків. Ситоподібні клітини
мають загострені кінці і скошені
поперечні перегородки. Ситоподібні
поля розташовані переважно на бічних
стінках. Деревина складається з
трахеїд, які мають облямовані пори і
паренхіми деревини, механічних
волокон немає. У серцевинних променях
часто накопичується смола, у цьому
випадку серцевинний промінь
розростається і перетворюється на
горизонтальний смоляний хід (рис. 82).
Горизонтальні смоляні ходи серцевинних
променів з'єднуються з вертикальними
смоляними ходами деревини, завдяки чому
створюється єдина видільна система.
Смоляні ходи вислані тонкостінними
парен- хімними клітинами.
За
вторинної будови стебло хвойних вкрите
перидермою, яка дуже швидко (4-5 років)
замінюється кіркою. У паренхімі первинної
кори є смоляні ходи, у флоемі немає
клітин-супутників, біля ситоподібних
клітин зустрічаються білкові клітини,
в яких міститься значна кількість
запасних білків. Ситоподібні клітини
мають загострені кінці і скошені
поперечні перегородки. Ситоподібні
поля розташовані переважно на бічних
стінках. Деревина складається з
трахеїд, які мають облямовані пори і
паренхіми деревини, механічних
волокон немає. У серцевинних променях
часто накопичується смола, у цьому
випадку серцевинний промінь
розростається і перетворюється на
горизонтальний смоляний хід (рис. 82).
Горизонтальні смоляні ходи серцевинних
променів з'єднуються з вертикальними
смоляними ходами деревини, завдяки чому
створюється єдина видільна система.
Смоляні ходи вислані тонкостінними
парен- хімними клітинами.
Ексцентричність деревини У природних умовах нахилені або зігнуті стовбури дерев і бічні гілки характеризуються нерівномірним розвитком шарів приросту на протилежних боках, що призводить до змі-

щення центру органа до одного боку. Такий нерівномірний розвиток річних кілець в осьових органах називають ексцентричністю деревини (рис. 83). Чим більшої деформації зазнає гілка або стовбур, тим виразніші ознаки ексцентричності. Внаслідок ексцентричної шаруватості річних кілець деревини розрізняють верхню, так звану тягову деревину, і нижню, креньову деревину, з різними біомеханічними властивостями, що протидіє вертикальній деформації під масою самої гілки чи її плодів.
Креньова деревина утворюється завдяки активнішій діяльності камбію на морфологічно нижньому боці гілок і пагонів, у результаті чого формується гіпоксилічна структура - значне переважання деревини на морфологічно нижньому боці органа. Така будова характерна для більшості представників голонасінних і деяких покритонасінних рослин.
Тягова деревина утворюється в результаті активнішого формування річ- них кілець на морфологічно верхньому боці пагонів і гілок більшості квіткових рослин, що теж супроводжується їхньою асиметричністю і створенням епіксилічної структури деревини.
Протилежні ділянки деревини відрізняються не тільки кількісно, а й якісно. Основою креньової деревини хвойних є великі, округлої форми трахеїди
з надзвичайно сильно потовщеними, твердими, повністю здерев'янілими клітинними стінками, що визначає високий рівень протидії стискуванню і згину цієї сторони. Анатомічні ознаки тягової деревини листяних рослин характеризуються великою кількістю волокон лібриформу і щільним їх з'єднанням, меншою кількістю судинних елементів. Це забезпечує високу міцність до розривів масивної верхньої сторони і протидію надмірним деформаціям гілок та крони.
Гігроскопічні зимові рухи гілок зумовлені тим, що товстіші клітинні стінки містять більше води, яку втрачають у морозні дні, а тому вони більше вкорочуються, а з ними вкорочується і відповідна ділянка деревини.
Будова стебла водних рослин
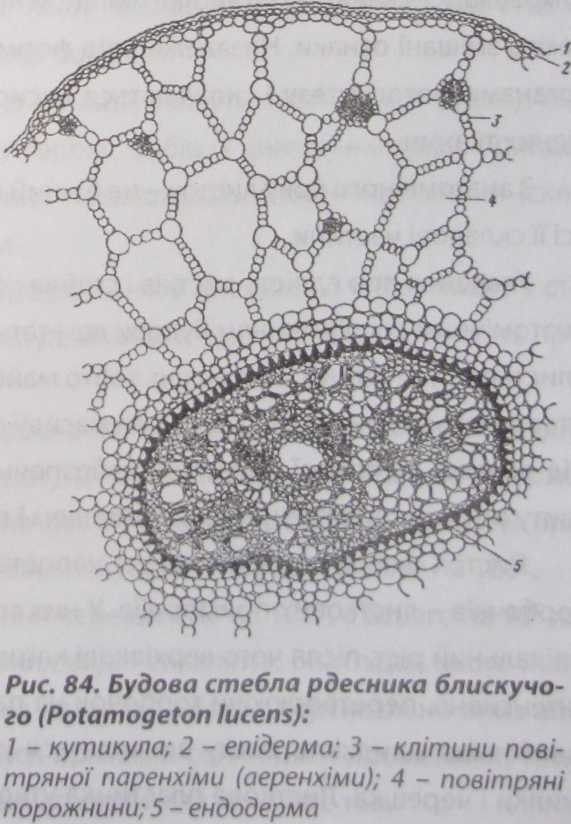 Особливості
водного середовища, насамперед висока
щільність води, нестача кисню та світла,
відбиваються на будові стебла водних
рослин (рис. 84). Епідерма занурених водних
рослин слабко диференційована, стінки
її клітин не потовщені, кутикула
редукована, клітини епідерми містять
хлоропласти, які беруть участь у
фотосинтезі, продихи відсутні.
Первинна кора займає більшу частину
поперечного зрізу стебла. Вона складається
із тонкостінної пухкої паренхіми, з
великими повітряними порожнинами, що
розташована одним або кількома
шарами, які відокремлені один від
одного тонкими перетинками - зазвичай
із одного шару клітин. Осьовий циліндр
має невеликі розміри, серцевина слабко
виражена, або взагалі відсутня. Провідні
пучки сильно зближені всередині
стебла, часто утворюється єдиний цен-
Особливості
водного середовища, насамперед висока
щільність води, нестача кисню та світла,
відбиваються на будові стебла водних
рослин (рис. 84). Епідерма занурених водних
рослин слабко диференційована, стінки
її клітин не потовщені, кутикула
редукована, клітини епідерми містять
хлоропласти, які беруть участь у
фотосинтезі, продихи відсутні.
Первинна кора займає більшу частину
поперечного зрізу стебла. Вона складається
із тонкостінної пухкої паренхіми, з
великими повітряними порожнинами, що
розташована одним або кількома
шарами, які відокремлені один від
одного тонкими перетинками - зазвичай
із одного шару клітин. Осьовий циліндр
має невеликі розміри, серцевина слабко
виражена, або взагалі відсутня. Провідні
пучки сильно зближені всередині
стебла, часто утворюється єдиний цен-
тральний тяж ксилеми, оточений флоемою. Ксилема містить невелику кількість судин, які у деяких рослин одразу після їх утворення руйнуються і тоді центр стебла займає вузький повітряний хід. У деяких видів, як, наприклад, у куширу зануреного (СегаІоіїНит детегзит), судини взагалі не утворюються. Здерев'яніння відбувається лише в стінках судин. Склеренхіма в повністю занурених стеблах не утворюється, механічна тканина тоді представлена виключно тяжами коленхіми. Камбій закладається в стеблах занурених рослин лише у деяких видів дводольних. Якщо закладається фелоген, то замість типової фелеми утворюється аеренхіма.