

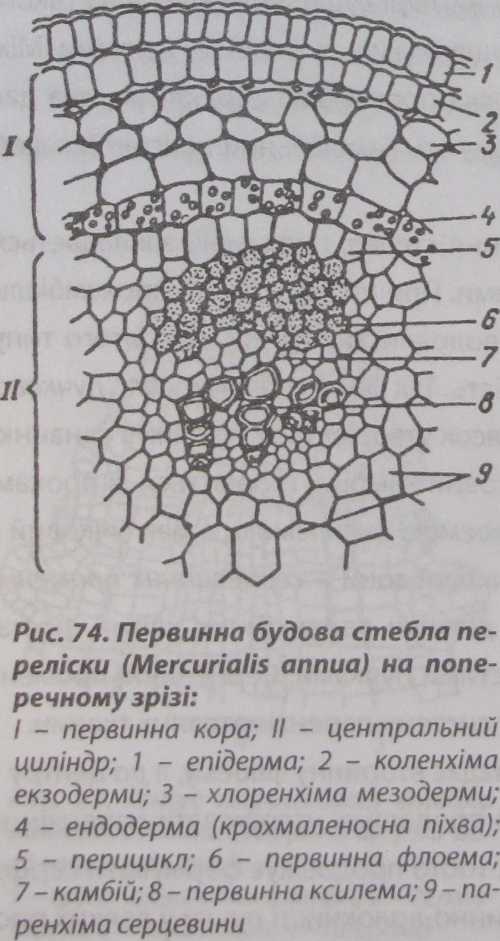
Первинна будова стебла
Насамперед на поверхні стебла утворюється епідерма, яка формується із дерматогену або туніки. Клітини її видовжені. У багатьох рослин на поверхні стебла зустрічаються волоски, які швидко осипаються.
Під епідермою диференціюється первинна кора, що складається із тонкостінних паренхімних хлорофілоносних клітин, завдяки чому молоді ділянки стебел мають зелене забарвлення. Периферійні шари клітин первинної кори часто представлені коленхімою, яка розташовується окремими зонами.
 У
внутрішніх шарах корової паренхіми
хлоропласти не зустрічаються. Цю
паренхіму називають основною. У
багаторічних рослин в ній відкладаються
запасні речовини.
У
внутрішніх шарах корової паренхіми
хлоропласти не зустрічаються. Цю
паренхіму називають основною. У
багаторічних рослин в ній відкладаються
запасні речовини.
У болотяних та водяних рослин основну паренхіму називають аеренхімою. Вона складається з мертвих тонкостінних клітин з великими повітряними порожнинами між ними. Така будова забезпечує ефективний газообмін.
Внутрішньою межею первинної кори є ендодерма. В стеблі з первинною будовою вона утворює крохма- леносну піхву - шар клітин, в яких містяться специфічні крохмальні зерна [статоліти), що беруть участь у геотропічних рухах (рис. 74). У стеблах типова ендодерма з поясками Каспа-
рі зустрічається рідко. Як правило це коренеподібні стебла (кореневища), які ростуть горизонтально на ґрунті або у верхніх шарах ґрунту (аїр).
За ендодермою до середини органа розташований осьовий, або центральний циліндр (стела). Він диференційований на перицикл (первинну латеральну меристему), зону провідних тканин та серцевину.
У стеблі з перициклу інколи утворюються волокна склеренхіми, які називають перициклічними.
Перицикл, як твірна тканина, може формувати комплекси клітин, з яких розвиваються адвентивні бруньки - бруньки, які розвиваються у незвичних місцях. Це має велике значення під час регенерації пошкоджених гілок, а також забезпечує вегетативне розмноження у природі.
У перециклі деяких рослин (кульбаба та ін.) утворюються членисті молочники, що містять смоли та латекс, а у зонтичних і звіробійних - секреторні канали, виповнені смолами та ефірними оліями.
Судинно-волокнисті пучки формуються із прокамбіальних тяжів, які є продовженням верхівкової меристеми. Спочатку із прокамбію формуються протофлоема і протоксилема. У протофлоемі містяться звичайні, але дрібні ситоподібні трубки. Протоксилема складається із спіральних та кільчастих трахеїд і трахей.
Терміни протофлоема і протоксилема застосовують до тих флоемних і ксилемних клітин, які ще здатні подовжуватися одночасно з видовженням органа. Частини флоеми і ксилеми, які виникають приблизно в момент завершення росту органа в довжину, називають метафлоемою і метаксилемою.
Протофлоема і метафлоема становлять первинну флоему, а протоксилема та метаксилема - первинну ксилему.
Центральну частину стебла займає серцевина. Вона складається з великих паренхімних клітин з міжклітинниками, які мають тонкі целюлозні стінки. У багатьох рослин у кінці першого періоду вегетації клітини серцевини відмирають, і клітинна порожнина заповнюється повітрям. У деяких багаторічників серцевина довгий час залишається живою і виконує функцію запасаючої тканини.
Якщо клітини серцевини відмирають одночасно, то її називають однорідною (бузина). У дуба, берези серцевина складається не тільки з живих, а й з мертвих клітин, тому її називають різнорідною.
Клітини серцевини рано досягають певного розвитку і їхній ріст при* пиняється. Якщо при цьому стебло продовжує потовщуватися, серцевина розривається, утворюючи порожнини, наприклад у зонтичних, губоцвітих та інших рослин.
Будова стебла однодольних рослин До класу однодольних належать трав'янисті (злакові, лілійні) і деякі деревні рослини (пальми).
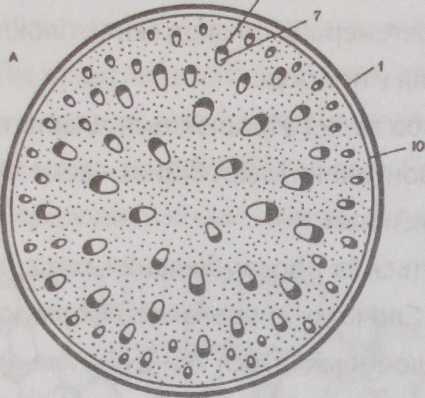 в Стебло однодольних має ряд
в Стебло однодольних має ряд
 особливостей
анатомічної будови: у них закриті
колатеральні судинно- волокнисті пучки,
немає камбію, справжнього вторинного
приросту, немає чіткої межі між
первинною корою і центральним циліндром
(рис. 75). У трав'янистих однодольних
рослин всі судинні пучки є листковими
слідами, тобто проходять від судинної
системи стебла в листки. Стебла мають
пучкову будову, коленхіма зустрічається
рідко. Фелоген у однодольних рослин не
утворюється. Немає ендодерми та
серцевини. Стебло не містить камбію, а
тому потовщується лише поки формуються
судинно-волокнисті пучки, в яких у
незавершеній формі зберігається
прокамбій. У зв'язку з відсутністю у
судинно-волокнистих пучках камбію
стебла однодольних мають первинну
будову впродовж усього життя рослини.
особливостей
анатомічної будови: у них закриті
колатеральні судинно- волокнисті пучки,
немає камбію, справжнього вторинного
приросту, немає чіткої межі між
первинною корою і центральним циліндром
(рис. 75). У трав'янистих однодольних
рослин всі судинні пучки є листковими
слідами, тобто проходять від судинної
системи стебла в листки. Стебла мають
пучкову будову, коленхіма зустрічається
рідко. Фелоген у однодольних рослин не
утворюється. Немає ендодерми та
серцевини. Стебло не містить камбію, а
тому потовщується лише поки формуються
судинно-волокнисті пучки, в яких у
незавершеній формі зберігається
прокамбій. У зв'язку з відсутністю у
судинно-волокнистих пучках камбію
стебла однодольних мають первинну
будову впродовж усього життя рослини.
Основна паренхіма стебла хлорофілоносна по периферії.
Рис.
76. Будова соломини жита (Зесаіе сегеаіе)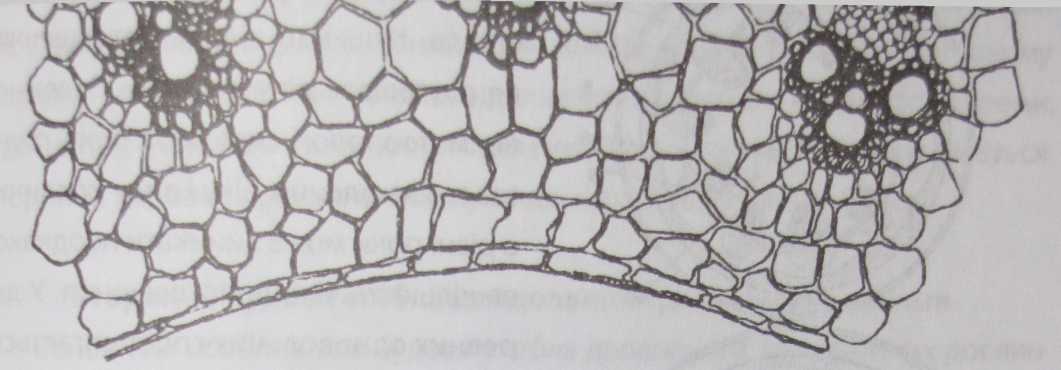
Колатеральні провідні пучки у злаків розташовані по колу периферії стебла у два ряди: внутрішній ряд складається з пучків більшого розміру. Провідні пучки мають механічні обкладки із склеренхіми (рис. 76).
Під епідермою розташована склеренхіма. Таке розташування склеренхіми надає міцності стеблам однодольних. Разом з тим, клітинні стінки епідерми та основної паренхіми товщають, дерев'яніють і мінералізуються. Це явище називають склерифікацією.
Потовщення стебла однодольних деревних рослин У деревних однодольних (пальми, алое та інші) за відсутності камбію спостерігається вторинне потовщення, яке відбувається завдяки формуванню особливого кільця потовщення. Кільце потовщення - це кільце, яке утворюється із клітин периферійної зони паренхіми первинної кори в результаті діяльності перициклічної групи клітин стебла. Клітини цього кільця активно діляться і відкладають до центру стебла нові меристематичні клітини, з яких формуються судинно-волокнисті пучки закритого типу: у пальм - колатеральні, у драцен - концентричні.
Одне кільце потовщення формує один ряд судинно-волокнистих пучків. Нове кільце потовщення з'являється за кільцем судинно- волокнистих пучків із паренхімних клітин периферійної частини. Подібні кільця потовщень виникають багаторазово і сприяють потовщенню стебла. Кількість кілець потовщення не відповідає віку рослини. Залежно від метеорологічних умов року, ґрунтового живлення, віку і виду рослини, в різні роки може виникати неоднакова кількість кілець потовщення. У деревних однодольних спостерігається лігніфікація міжпучкової паренхіми. Паралельно з формуванням кілець потовщення епідерма замінюється перидермою.
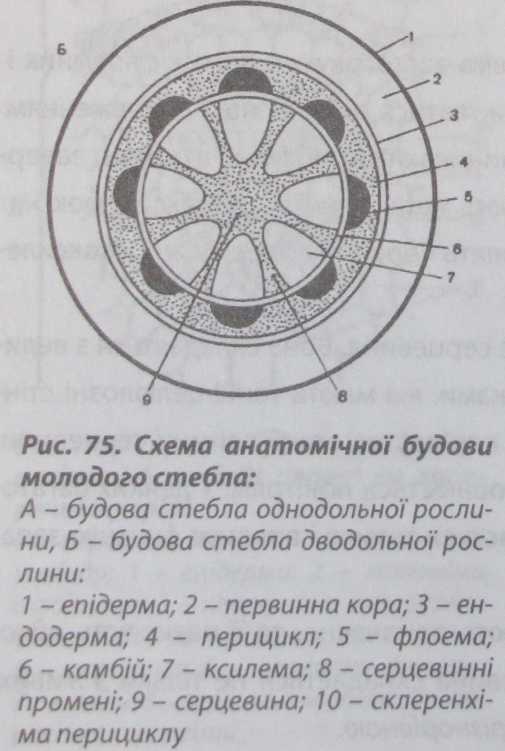
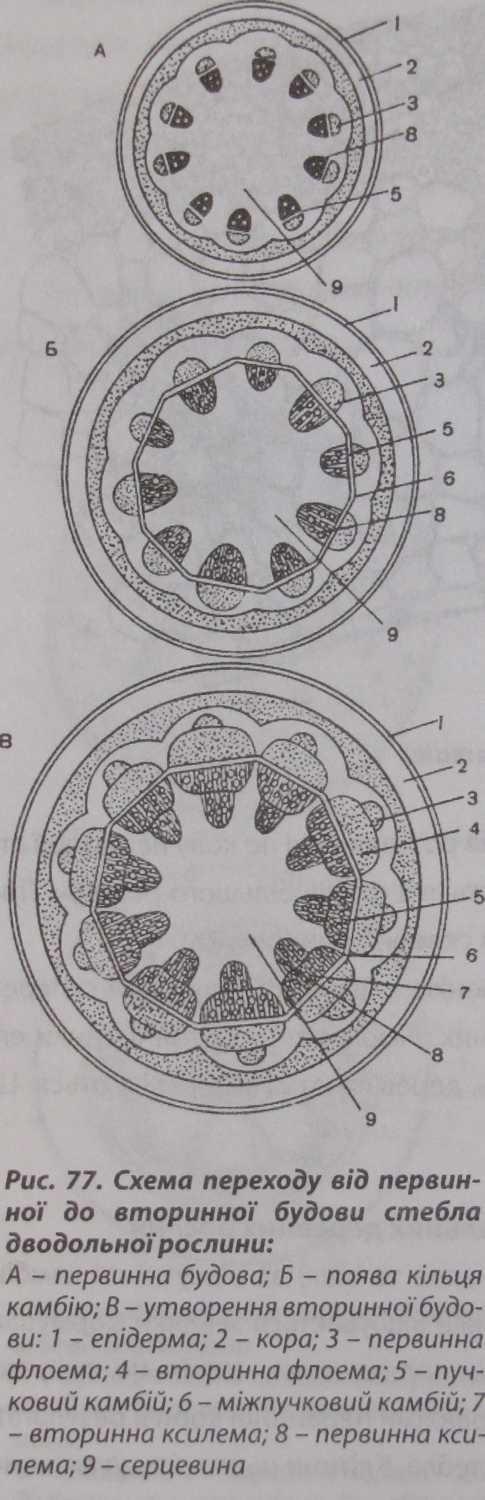 Будова
стебла дводольних рослин Вторинні
меристеми у стеблах дводольних рослин
диференціюються рано, утворюючи
вторинні структури. Вторинні зміни
у будові стебла дводольної рослини
починаються з диференціації камбію
(рис. 77). Цей процес у деревних порід
завершується настільки швидко, що у
пагона, який тільки розпустився дуже
важко розрізнити структурні елементи
первинного походження.
Будова
стебла дводольних рослин Вторинні
меристеми у стеблах дводольних рослин
диференціюються рано, утворюючи
вторинні структури. Вторинні зміни
у будові стебла дводольної рослини
починаються з диференціації камбію
(рис. 77). Цей процес у деревних порід
завершується настільки швидко, що у
пагона, який тільки розпустився дуже
важко розрізнити структурні елементи
первинного походження.
У трав'янистих рослин, незважаючи на раннє формування вторинних утворень, гістологічні елементи первинної будови зберігаються довго.
У трав'янистих рослин типову первинну будову стебла можна спостерігати у бруньці, яка готова до розпускання.
На початку розпускання бруньки з'являється камбій, який активно функціонує на перших етапах і формує вторинні елементи. Камбій диференціюється із міжпучкових паренхімних клітин і частково із прокамбіальних клітин, які збереглися між флоемою і ксилемою. З активних прокамбіальних клітин формується пучковий камбій, а з активованих прошарків міжпучко- вої паренхіми - міжпучковий камбій. Згодом пучковий і міжпучковий камбії, з'єднуючись, утворюють камбіальне кільце. Камбій функціонує у такому порядку: назовні клітини камбію відкладають елементи вторинної флоеми, а до центру - елементи вторинної ксилеми. Одночасно місцями утворюється вторинна корова паренхіма, формуються вторинні луб'яні волокна.