

Анатомічна будова стебла
Анатомічна будова стебла повністю відповідає тим фізіологічним функціям, які воно виконує: опорній, провідній, асиміляційній та запасаючій.
Структура стебла формується в результаті діяльності його верхівкової (апікальної) меоистеми (синоніми апекс, конус наростання і тгш«*
У квіткових та голонасінних рослин верхівкова меристема має вигляд конуса з округлою верхівкою, який складається з групи меристематичних клітин, і розташована в основі бруньки (рис. 72).
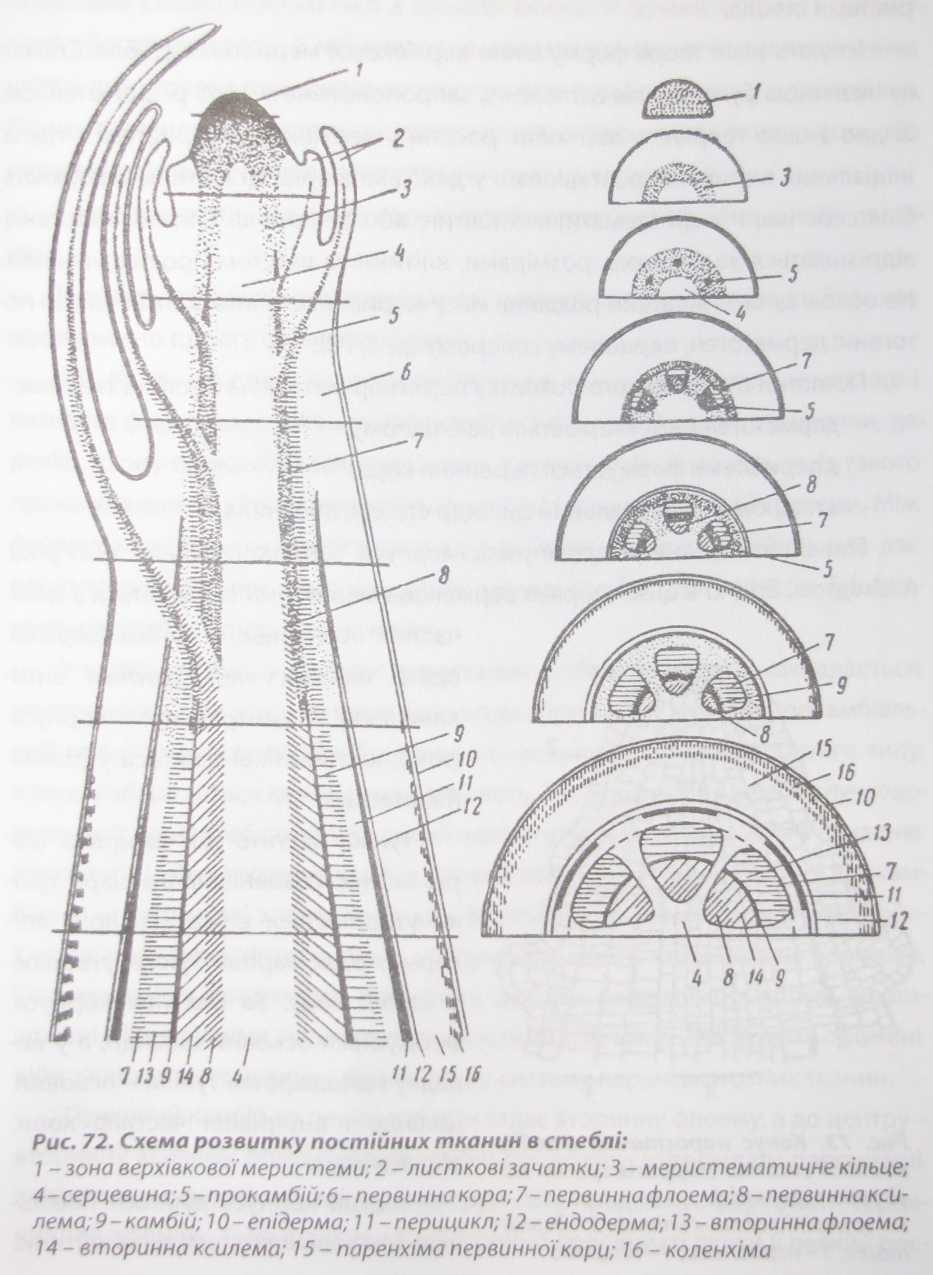
Верхівкова меристема - найактивніша та важлива частина пагона, його ростовий центр. Вона формує прокамбіальні тяжі, листки, вегетативні та генеративні органи. Листки і бруньки теж беруть початок від верхівкової ме- ристеми стебла.
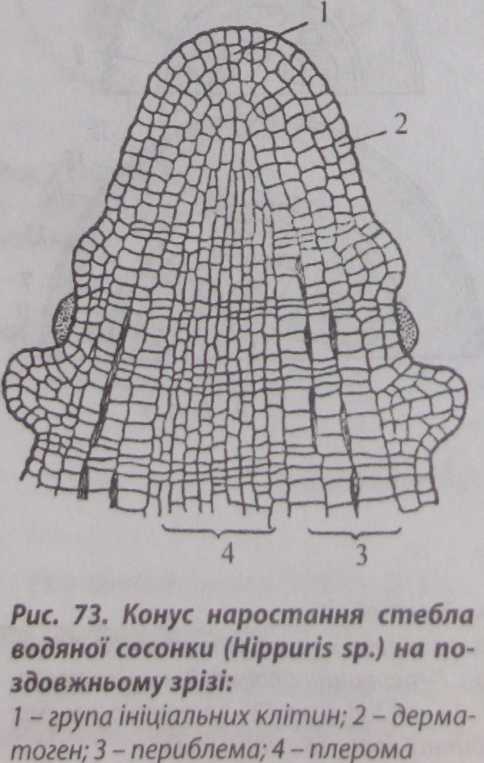
Існують різні теорії формування верхівкової меристеми стебла. Спочатку визнаною була теорія гістогенів, запропонована у 1868 р. І.Ганштейном. Згідно з цією теорією у квіткових рослин у верхівковій меристемі є група ініціальних клітин, які розташовані у декілька шарів. Ці клітини утворюють біля себе шари меристематичних клітин, або гістогени. Клітини гістогенів відрізняються за формою, розмірами, клітинним вмістом і розташуванням. На основі цього Ганштейн розділив масу меристематичних клітин на три гістогени: дерматоген, периблему і плерому (рис. 73).
Гістогени в міру їхнього розвитку перетворюються на постійні тканини:
дерматоген перетворюється на епідерму;
з периблеми формується первинна кора;
| з плероми і центральний циліндр стебла, або стела.
Більш визнаною є теорія туніки і корпуса, запропонована у 1924 році А.Шмідтом. Згідно з цією теорією верхівкова меристема складається з двох
 частин:
зовнішньої І
туніки
(верхній одяг), клітини якої діляться
антиклінально та центральної - корпуса
(тіло), клітини якої діляться у різних
напрямках.
частин:
зовнішньої І
туніки
(верхній одяг), клітини якої діляться
антиклінально та центральної - корпуса
(тіло), клітини якої діляться у різних
напрямках.
Туніка містить до чотирьох шарів клітин. Із зовнішнього шару туніки утворюється епідерма. Крім епідерми, одношарова туніка утворює в стеблі кору. За рахунок корпуса формується осьовий циліндр, а у випадку одношарової туніки - осьовий циліндр і внутрішня частина кори, або вся кора.
Ініціалі корпуса діляться повільніше ніж інші меристематичні клітини.
Швидкість поділів зростає, коли апікальна меристема отримує флоральний стимул. Тому ці ініціалі назвали меристемою очікування.
За допомогою сучасних методів дослідження показано, що верхівкова меристема стебла складається з більшої кількості функціонально і цитологічно обособлених зон. У ній виявлено зону центральних меристематичних клітин, яка оточена периферійною (органогенною, фланговою) меристемою. Під центральною меристемою відокремлюється стрижнева (колончаста, або серцевинна) меристема, яка перетворюється на серцевину.
Слід зауважити, що поділ верхівкової меристеми на зони досить умовний.
В результаті діяльності верхівкової меристеми формується стебло, яке має відносно просту первинну будову.
С.П. Костичев довів, що прокамбій може виникати в двох формах, що і визначає формування різних типів стебла. У більшості деревних рослин, деревію, льону прокамбій закладається у вигляді суцільного кільця. Із такого прокамбіального кільця формуються суцільні кільця флоеми і ксилеми. Між флоемою і ксилемою зберігається смужка прокамбіальних клітин, яка дає початок камбіальним клітинам, що формують камбіальний поясок. Так формується стебло безпучкового типу.
У інших рослин, у конусах наростання стебел, прокамбій закладається не суцільним кільцем, а окремими тяжами. При цьому кожний прокамбіаль- ний тяж формує самостійний судинно-волокнистий пучок відкритого типу, в якому зберігається камбіальна діяльність. Так формується стебло пучкового типу. У таких стеблах камбіальний поясок утворюється завдяки з'єднанню пучкового і міжпучкового камбіїв. Пучковий камбій утворюється із прокамбіальних клітин, які знаходяться між флоемою і ксилемою, а міжпучковий - з прошарку паренхімних клітин міжпучкової зони - серцевинних променів. Серцевинними променями називають ділянки паренхімних клітин, розташовані між сусідніми судинно-волокнистими пучками. Серцевинні промені ніби з'єднують серцевину і кору в єдину систему паренхіматозних тканин.
Пучковий камбій до периферії відкладає вторинну флоему, а до центру - вторинну ксилему. Міжпучковий камбій продовжує утворювати паренхімні клітини як до периферії, так і до центру, тобто продовжує формувати серцевинний промінь. Тому індивідуальні судинно-волокнисті пучки у деяких рос
лин не можуть зблизитись і залишаються розмежованими протягом усього життя рослини.
Пучковий тип будови стебла здебільшого зустрічається у дводольних трав'янистих рослин, наприклад у жовтецю, соняшнику, маку городнього, а з деревних рослин - саксаулу і винограду.