

Хромосомы человека
История развития цитогенетики человека
Впервые митотические хромосомы человека были описаны в работах Дж. Арнольда (1879) и В. Флемминга (1882). В последующие годы различные оценки их количества давали результаты от 47 до 49 хромосом, причем у мужчин и женщин находили разное их число. Эти первые исследования проводились на гистологических срезах тестикул или яичников. В то время техника получения срезов была такова, что митозы в готовых препаратах были, как правило, разрушены. Хромосомы на них накладывались одна на другую, образовывали клубки и плохо поддавались анализу. Эти трудности удалось преодолеть только к 50-м гг. XX в., когда для получения препаратов хромосом стали использовать суспензии клеток, выращенных в клеточных культурах. Так, первые препараты хромосом человека с хорошим разрешением были получены на клетках фибробластов эмбриона легкого, выращенных in vitro. Суспензии клеток промывали гипотоническим раствором, в результате чего клетки набухали и лопались, а хромосомы свободно распределялись на стекле. Позже этот способ был усовершенствован. Перед гипотоническим шоком на клетки воздействовали колхицином — веществом, которое, разрушая нити веретена деления, останавливает митоз на стадии метафазы, когда хромосомы наиболее легко идентифицировать. Это позволило получить большое число клеток в метафазе митоза, и препараты, приготовленные таким образом были более удобны для подсчета хромосом. Пользуясь подобным методическим подходом, в 1955 году А. Леван и Дж. Тио, изучив 261 метафазную пластинку, пришли к выводу, что количество хромосом в клетках человека равно 46, причем как в мужских, так и в женских клетках. Годом позже и другие исследователи на препаратах тестикул трех пожилых мужчин на стадии метафазы мейоза 1 нашли 23 бивалента, что соответствует 46 хромосомам в диплоидном наборе. Эти результаты ознаменовали возникновение новой отрасли исследований — клинической цитогенетики. В настоящее время цитогенетика человека достигла высокого уровня и находится на переднем крае фундаментальной цитогенетики.
Нормальный кариотип человека
Препараты хромосом человека можно приготовить из любых тканей и клеточных суспензий, но лишь, если в них содержатся делящиеся клетки, т. к. вне деления (во время интерфазы) хромосомы деспирализуются и переходят в состояние хроматина Чаще всего препараты готовят из клеток костного мозга, кратковременной культуры клеток крови или из перевиваемой культуры фибробластов. Наиболее прост и доступен метод культивирования клеток крови лейкоцитов и лимфоцитов. Митозы в таких культурах стимулируют искусственно. Чтобы остановить жизненный цикл клеток в прометафазе, в них подавляют образование веретена деления, обрабатывая веществами с колхициноподобными свойствами. Для свободного распределения хромосом на стекле клетки обрабатывают гипотоническим раствором, вызывая гипотонический шок. Затем препарат фиксируют смесью этанола и уксусной кислоты, высушивают и окрашивают. Когда с помощью стандартных методов хромосомы окрашиваются целиком, равномерно и интенсивно, их систематизируют согласно Денверской классификации, принятой в 1960 г., нумеруя пары хромосом от 1 до 23. В кариотипе человека различают метацентрические, субметацентрические и акроцентрические хромосомы. Учитывая относительную длину плечей, положение центромеры и центромерный индекс, который отражает процентное соотношение длин короткого плеча и всей хромосомы, 23 пары хромосом человека разбивают на 7 групп (рис .13). В группу А (№№ 1-3) входят пары наиболее крупных метацентрических аутосом. Группа В (№№ 4-5) объединяет две пары субметацентрических хромосом, неразличимых между собой. Группа С (№№ 6-12) содержит семь пар аутосом среднего размера. Размеры и форма этих хромосом неодинаковы, однако стандартные методы окрашивания не позволяют их идентифицировать. В группу D (№№ 13-15) объединены три пары акроцентрических хромосом среднего размера, морфологически сходных между собой. Все хромосомы группы D содержат спутник, который не всегда выявляется, может быть очень большим, а иногда и двойным. Длина короткого плеча этих хромосом также изменчива. К группе Е (№№ 16-18) относятся три пары почти метацентрических хромосом, из которых в 16-й паре центромера наиболее близка к середине, а две другие пары неотличимы друг от друга. Группа F содержит мелкие метацентрические аутосомы (№№ 19-20), группа G — мелкие акроцентрические (№№ 21-22). Внутри групп F и G нары хромосом неразличимы. Длина коротких плечей у них изменчива, как и у хромосом группы D. Короткие плечи хромосом групп D и G содержат районы ядрышкового организатора. Перечисленные 22 пары хромосом относятся к аутосомам, одинаковым у мужчин и женщин. Половые хромосомы составляют 23-ю пару. У женщин — это две Х-хромосомы. У мужчин — Х- и Y-хромосомы. Половая Х-хромосома неотличима от аутосом группы С. При стандартном окрашивании она включается в состав этой группы: №№ 6-12 и X. Мужская половая Y-хромосома является акроцентрической, сходна по морфологии с хромосомами группы G, но ее легко отличить по морфологическим критериям. Длина короткого плеча Y-хромосомы изменчива и индивидуальна, причем варианты длины плеча наследуются от отца к сыну. Y-хромосома, в отличие от хромосом последней группы, не имеет спутников. В интерфазных ядрах концевой участок длинного плеча Y-хромосомы можно выявить в составе хроматина, пользуясь специфическим окрашиванием акрихин-ипритом : в результате этот участок выявляется как яркое пятно диаметром 0,3-1,0 мкм,
Во многих хромосомах человека обнаружены ломкие (фрагильные) участки, подверженные хромосомным и хроматидным разрывам. Такие разрывы легко получить в клеточных культурах, удаляя фолиевую кислоту из питательной среды культивируемых клеток, В настоящее время показано, что одна из форм умственной отсталости человека связана с наличием определенного фрагильного участка в концевом районе длинного плеча Х-хромосомы.
Длина одной и той же хромосомы в разных фазах митоза различна, поскольку конденсация хромосом продолжается до конца метафазы. Кроме того их конденсация значительно усиливается в присутствии применяемого для приготовления препаратов колхицина.
Анализ препаратов хромосом человека показал, что в ряде случаев, как уже говорилось выше, на некоторых хромосомах могут существовать вторичные перетяжки. Спутничными перетяжками обладают все акроцентрические хромосомы (нары №№ 13, 14, 15,21,22). Вторичная перетяжка бывает также в аутосомах пары № 9. В них она располагается в околоцентромерном районе длинного плеча.
Дифференциальное окрашивание хромосом
Современные цитогенетические методики позволяют идентифицировать по морфологии все пары хромосом на препарате, а в ряде случаев и хромосомы внутри одной пары. Суть этих методик состоит в дифференциальном окрашивании нативных хромосом по длине, что обеспечивается сравнительно
простыми температурно-солевыми воздействиями на фиксированные хромосомы или использованием специфических красителей (рис. 3.19). Дифференциальное окрашивание приводит к появлению линейного рисунка но длине хромосомы.
Несмотря на большое разнообразие способов обработки хромосомных препаратов и красителей, выявляемый линейный рисунок хромосомы всегда один и тот же. Он меняется только в зависимости от степени конденсированное™ хромосомы. Сегмент, видимый как одна полоса в метафазной хромосоме, в менее конденсированной прометафазной хромосоме, может предстать в виде нескольких мелких полос.
Дифференциальное окрашивание в зависимости от используемого метода может охватывать либо всю длину хромосомы, либо ее центромерный район.
Представление о рисунке дифференциально окрашенных по всей длине хромосом можно получить, окрашивая препараты по G-методу с использованием красителя Гимзы. В этом случае хромосомы выглядят состоящими из поперечноисчерченных, по-разному окрашенных сегментов. Каждой паре хромосом присущ индивидуальный рисунок исчерченности за счет неодинаковых размеров сегментов. В мелких хромосомах рисунок образуется единичными сегментами, в крупных хромосомах сегментов много
Общее для нормального хромосомного набора число окрашенных и неокрашенных сегментов в метафазе составляет около 400. В прометафазных хромосомах оно увеличивается до 850 и более.

Разные типы сегментов получили обозначения G-, Q-, R-, С-, и Т по названиям методов, с помощью которых они выявляются. Так, для G-сегментов это метод Гимза после предварительной обработки фиксированных хромосом. Q-сегменты после окрашивания акрихинипритом приобретают яркое флюоресцирующее свечение. Рисунок Q и G-сегментов полностью идентичен. R-сегменты расположены между Q-сегментами.
Место нахождения С-сегментов можно обнаружить с использованием других методов окрашивания. Эти сегменты связаны с локализацией конститутивного, или структурного гетерохроматина. В разных хромосомах размер С-сегментов неодинаков. Гетерохроматин, обнаруженный по методу С-окраски, содержится во всех хромосомах человека. Он находится в околоцентромерных районах всех хромосом, а также в дистальной части длинного плеча Y-хромосомы, на длинных плечах 1-й, 9-й и 16-й пар хромосом
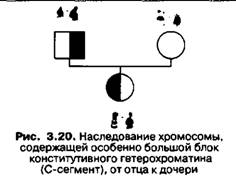
Небольшие блоки структурного гетерохроматина выявляются в плечах 2-й нары и Х- хромосом. В акроцентрических хромосомах гетерохроматин С-сегментов расположен в коротких плечах. Накопленные сведения говорят, что в разных популяциях человека размеры сегментов гетерохроматина значительно различаются. Морфологические особенности сегментов наследуются по законам Менделя (рис. 3.20). Особенно изменчива величина С-сегментов в аутосомах №№ 1, 4, 9, 13-15, 16, 21-22 и Y-хромосоме. Поскольку у большинства носителей таких особенностей кариотипа отсутствуют фенотипические аномалии, то подобные особенности в строении хромосом можно рассматривать как варианты нормы. Согласно данным общей цитогенетики, значительное количественное изменение гетерохроматина в кариотипе не оказывает сильного отрицательного влияния на развитие организма человека.
Еще один тип сегментов — Т — выявляется также с помощью специфических методов окрашивания и присутствует в теломерных районах всех хромосом.
В метафазе митоза может быть выявлена еще одна характеристика линейной неоднородности хромосом. Она связана с асинхронностью репликации ДНК в их разных участках. Такой тип окрашивания разбирается подробно в главе 4 в разделе, посвященном клеточному циклу.
Индивидуальная совокупность сегментов, различающихся по ширине и интенсивности окрашивания, образует цитологическую карту каждой хромосомы. Основанные на дифференциальном окрашивании хромосом цитологические карты имеют исключительное значение для развития цитогенетики человека. С помощью этих карт стало реальным выяснить происхождение аномальных хромосом, вплоть до точного описания, какие конкретно районы вовлекаются в ту или иную форму хромосомного нарушения. На международных совещаниях по номенклатуре в цитогенетике человека была разработана и введена в практику система обозначения сегментов нормальных хромосом и хромосом, подвергшихся тем или иным структурным перестройкам.
Дальнейшее совершенствование методов окрашивания хромосом позволило выявить до 1000 полос на всех 23-х хромосомах человека, характерных для гаплоидного набора. В среднем на хромосому при этом приходится 50 полос, хотя на некоторых хромосомах их можно обнаружить в несколько раз больше, чем на других. Гаплоидный геном человека состоит из 3x109 пар нуклеотидов, соответственно каждая полоса содержит в среднем 3x106 нар нуклеотидов, что соответствует нескольким сотням генов.
Половой гетерохроматин
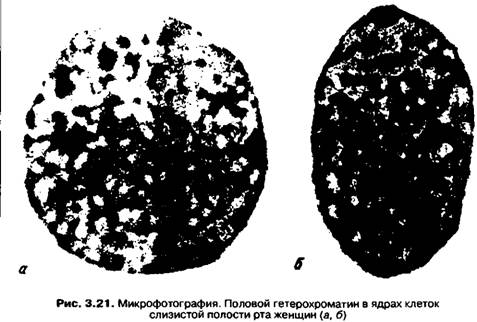
В соматических клетках женщин половой хроматин выявляется в виде гетерохроматина - небольшой хорошо окрашенной округлой структуры, размером 0,8-1,1 мкм, находящейся возле ядерной мембраны (рис. 3.21). Половой хроматин называют также тельцем Барра, т. к. впервые он был описан этим ученым в нейронах кошки. Позже оказалось, что половой гетерохроматин присутствует в соматических клетках всех млекопитающих женского пола, в том числе и человека. Гомологичные половому хроматину структуры, так называемые "барабанные палочки" были обнаружены в ядрах полиморфноядерных лейкоцитов. Половой гетерохроматин — это одна из Х-хромосом, которая находится в неактивном, суперснирализованном состоянии. Известно, что фенотипически пол у человека определяется наличием или отсутствием Y-хромосомы, а не количеством Х-хромосом. Если в кариотипе зиготы присутствует хотя бы одна Y-хромосома, а количество Х-хромосом превышает единицу, то по фенотипу формируется мужчина
Количество телец Барра в клетках всегда на одно меньше, чем число Х-хромосом. То есть только одна Х-хромосома в соматических клетках человека, и мужчины, и женщины, всегда находится в активном состоянии. В норме женщина имеет две, а мужчина одну Х-хромосому, в связи с чем инактивация второй Х-хромосомы у женщин в виде полового гетерохроматина служит механизмом компенсации различий в дозе генов, не оказывающих влияния на развитие половых признаков и признаков, сцепленных с Х-хромосомой. Этот же механизм оказался фактором, благоприятствующим носителям Х-хромосомных анеуплоидий. Какое бы количество Х-хромосом они не несли, генетически активна только одна. Остальные же Х-хромосомы существуют в виде факультативного полового гетерохроматина. Поэтому но количеству телец Барра в соматических клетках можно диагносцировать форму анеуплоидий. Например, у женщин с кариотином 47, XXX обнаруживаются два тельца Барра, а с кариотипом 45, ХО — ни одного. У мужчин с кариотипом XXY — одно.
Образование полового хроматина из Х-хромосомы происходит на ранних стадиях эмбрионального развития. Дробящаяся "женская" зигота млекопитающих имеет две функционально активные Х-хромосомы. У человека половой хроматин появляется на стадии развития зародыша в несколько сотен клеток. В трофобласте Х-хроматин выявлен на 12-й день развития, а в собственно эмбрионе — на 16-й день. Половой хроматин образуется сразу во всех клетках эмбриона. В настоящее время наиболее приемлемой является гипотеза, согласно которой в разных клетках одного организма могут быть инактивнрованы разные Х-хромосомы: в одних — отцовская, в других — материнская. То есть по Х-хромосоме женщины мозаичны. Это положение можно рассмотреть на примере активности фермента глюкозо-6-фосфат дегидрогеназы (Г6ФД), ген которого находится в Х-хромосоме. Женщины имеют два аллеля этого гена, а мужчины только один. Тем не менее средний уровень активности этого фермента сходен у представителей обоих полов, следовательно, должен действовать механизм дозовой компенсации. У гетерозиготных женщин с помощью электрофореза в разных клетках можно выявить два аллеля Г6ФД. В ряде работ на эритроцитах in vivo и на фибробластах соединительной ткани в клеточных культурах у женщин был выявлен мозаицизм по Г6ФД.
Тельца Барра присутствуют не во всех клетках женщины. Так, обе Х-хромосомы активны в ооцитах и в клетках женской половой системы.