

2.2. Биономический анализ
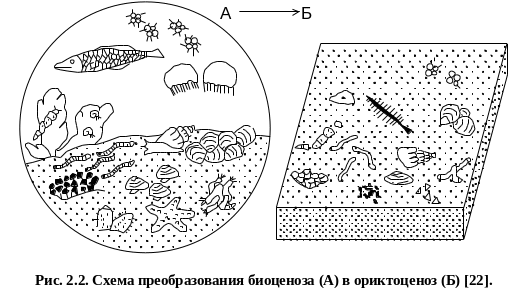
Биономический анализ – это изучение ориктоценозов с целью восстановления условий обитания организмов и палеогеографических обстановок. Ориктоценоз («ископаемое сообщество») это сохранившаяся в ископаемом состоянии группировка самых различных по происхождению остатков организмов. Путь исследования – воссоздание палеобиоценозов (сокращенно палеоценозов), изучение остатков организмов, восстановление их образа жизни и среды обитания, в конечном итоге – палеогеографии. Решение этой задачи осложняется тем, что в ряду палеоценоз – танатоценоз (совокупность в каком либо месте остатков организмов, живших здесь и привнесенных сюда из других мест) – тафоценоз (остатки перемещенных организмов) – ориктоценоз происходит существенное изменение комплекса остатков организмов. Поэтому ориктоценоз может являться скоплением небольшой части организмов, некогда входивших в различные палеоценозы (рис. 2.2).
При биономическом анализе следует идти в обратном направлении – анализируя ориктонозы, восстанавливать облик отдельных организмов и затем переходить к реконструкциям древней географической обстановки с палеоценозами. Прежде всего, изучают остатки отдельных организмов, входивших в палеобиоценоз. Принадлежность остатков к той или иной система-тической группе указывает на определенную обстановку, так как каждая группа организмов обитала в определенных условиях. Затем переходят к изучению экологических групп организмов и, наконец, к палеобиоценозу (палеоценозу) в целом. Видовой состав сообщества свидетельствует о нормальной (виды разнообразны, количество особей может быть незначительным) или о специфической (видов мало, но большое количество особей) обстановках обитания.
Ценные сведения можно получить при изучении формы тела или его положения, особенно у сидячего бентоса, а также размеров тела, толщины створок раковины. Не менее важную информацию даёт исследование следов жизнедеятельности организмов (палеоихнология): следы ползания по дну, зарывания в донные осадки, сверления в твердом субстрате и т. д. Всё это указывает на тип среды (водная, воздушная), газовый режим, динамику среды и характер грунта.
Особенности захоронения позволяют делать выводы о движениях водной среды. Скопление ломаной, окатанной или целой ракуши свидетельствует о переносе течениями или волнами, ори-ентировка в пространстве удлиненных раковин указывает направление движения воды, остатки скелетов с сохранившимися тонкими деталями – о спокойной водной среде осадконакопления.
Биономический анализ заключается в определении фаций преимущественно на основе изучения органических остатков, следов жизнедеятельности организмов. Для этого надо иметь представление об условиях жизни растений и животных, об основных факторах, определяющих их распространение и развитие. Этому помогает палеоэкология, выясняющая взаимоотношения когда-то существовавших организмов с окружающей их средой, состоящей из абиотических и биотических факторов. Биос и окружающая среда образуют экосистему, представляющую собой влияющие друг на друга взаимосвязанные компоненты. На образ жизни различных организмов влияет ряд факторов.
Глубина бассейнов определяет освещённость давление (оно через каждые 10 м глубины увеличивается на 105 Па), смену газового режима. Для жизни животных и растений наиболее благоприятны небольшие глубины. С их увеличением число видов и количество биомассы значительно уменьшается. На больших глубинах она в 100-200 тысяч меньше, чем на шельфе, где сосредоточено около 83% всей биомассы бентоса. Определить глубину древних бассейнов сложно и возможно только по остаткам бентосных организмов. На небольших глубинах обитали водные растения и разнообразные рифостроители: известьвыделяющие водоросли (строматолиты), археоциаты, кораллы, строматопораты. Массовые поселения брахиопод и двустворок характерны для морского дна неглубокого бассейна. На мелководье располагались брахиоподовые и пелециподовые банки, заросли морских лилий.
Относительное мелководье или шельф – зоны литорали и неритовая, сравнительное мелководье – эпиконтинентальные бассейны. Глубоководные области – континентальный склон (батиаль) и дно океана – абиссаль.
Солёность бассейнов. Измеряется в промилле (1%=1г/л). Солёность Мирового океана составляет 35%, открытых морей близка к этому значению, внутренних бассейнов, или глубоко вдающихся в континент, отличается. В Красном море она 43%, Каспийском – 17, Балтийском – до 9. В пресноводных бассейнах солёность ниже 0,5%, солоноватоводных – 5-15, осолонённых – свыше 45%. Солёность древних бассейнов также колебалась, состав солей в них менялся. Наиболее богатая и разнообразная фауна обитает в морях нормальной солёности. Так в Средиземном море число видов превышает 8000, а в Азовском (16%) сокращается в 15-20 раз. В бассейнах с ненормальной солёностью условия благоприятны для немногих видов, однако при этом может резко возрастать их продуктивность. Для таких бассейнов часто характерны бедность видами и богатство особями. Изменение солености приводит к «угнетенному» облику фауны: уменьшаются размеры раковин (например, двустворка Cardium edule в Кильской губе имеет длину 44 мм, а Ботническом заливе всего 18 мм), они становятся тоньше, упрощается их скульптура.
Выделяются виды эвригалинные и стеногалинные. Первые выдерживают колебания солёности, вторые могут жить только в водах нормальной солёности. К стеногалинным формам относятся колониальные кораллы, иглокожие, головоногие моллюски, брахиоподы, трилобиты. Только единичные представители перечисленных групп могли переносить изменение солености. Эвригалинные – это пелецеподы, мшанки, ракообразные, черви, водоросли, бактерии. Наиболее выносливы последние четыре группы. Однако представители некоторых из перечисленных групп не выносят изменения солености. Так, двустворки Hippurites, Diceras жили в морях нормальной солености. По органическим остаткам выясняют, в каком по солёности бассейне обитали животные.
Свет необходим для фотосинтеза растений. Наиболее освещены верхние 10 м водной толщи, а он проникает обычно до глубин 80м. По освещённости выделяется эвфотическая (поверхность-30-80 м), дисфотическая (30-80-200 м) и афотическая (глубже 80-200 м). На освещённых глубинах моря наиболее богатый растительный, а, следовательно, и животный миры, а ниже отметки 200 м царит полный мрак. Развитие растительности в освещенных верхних слоях воды (до глубины 50-80 м) и на мелководье приводит к тому, что здесь существует наиболее богатый животный мир, представленный растительноядными формами, разнообразными хищниками, трупоедами и илоедами. Проникновение солнечного света на глубину зависит от прозрачности воды и географической широты местности. В Средиземном море белый диск, опущенный в воду, виден до глубины 60 м, в Белом – до 8-9 м, в Азовском – до 3 м, а в летнее время из-за массового развития одноклеточных водорослей снижается до 10-12 см.
Температура воды определяется географической широтой, временем года, глубиной бассейна. Вода отличается большей термостабильностью, чем воздух, у неё низкая теплопроводность. В бассейнах существует «температурная слоистость». Так, зимой холодные воды подо льдом располагаются на более теплых, а летом более легкие прогретые воды находятся вверху. Такие условия благоприятны для обитателей водной среды. Температура воды в Мировом океане обычно колеблется от -2 до 20 °С, наивысшая – 36 °С. Все глубинные области заполнены холодными водами.
Относительные значения температур могут быть определены по биоценозам животных и растительных сообществ. Температурный фактор сильно сказывается на жизнедеятельности литоральных бентосных и планктонных форм. Он определяет саму возможность существования организмов, вызывает изменение размеров и формы тела, регулирует темп роста и влияет на продолжительность жизни. Наиболее восприимчивы к температурному режиму планктонные водоросли, среди которых особенно важны кокколитофориды и диатомовые. Важные сведения о температурах морских вод и прилегающих континентов дает анализ споро-пыльцевых комплексов, захороненных в прибрежных осадках.
Выделяют эвритермные и стенотермные организмы. Первые переносят колебания температуры, вторые – нет. Так, колониальные кораллы не переносят температуры ниже 20 °С. Наиболее богатая и разнообразная фауна в тропических морях. В них раковины животных более толстые, массивные, с богатой скульптурой. В холодной воде растворимость карбоната увеличивается, поэтому осадки морей полярных и умеренных широт обеднены им, а раковины обитающих здесь животных тонкие и с простой скульптурой. Температура воды влияет на растворимость фосфатов, большую в холодных водах. В зонах апвеллинга (выходов холодных вод, несущих фосфор, на поверхность) развита бурная жизнь. Органика, попадающая в осадок после отмирания организмов и растений, фоссилизируется и превращается затем в фосфориты.
Газовый режим определяется содержанием О2, Н2S, СО2 и имеет большое значение для водного населения. Кислород поступает из атмосферы и выделяется водными растениями. Он необходим для нормальной жизнедеятельности организмов. Содержание кислорода, необходимого организмам для дыхания, зависит от температуры и характера циркуляции вод. Проникновение этого газа в глубокие части бассейнов зависит от вертикального распределения плотности и солености вод, перемещения водных масс. В водах полярных морей кислорода почти в два раза больше, чем в тропических.
Углекислота выделяется в воду организмами и поступает в неё во время вулканической деятельности. Углерод потребляется фото- и хемосинтзирующими растениями и расходуется на химические соединения. С повышением температуры и солёности содержание СО2 снижается. В высоких концентрациях он ядовит. На ненормальный газовый режим указывает присутствие в осадках большого количества мелких кристаллов пирита или сидерита, приуроченных к плоскостям напластования.
Сероводород образуется в результате деятельности бактерий и для водных животных смертелен. Пример – Чёрное море с сероводородным заражением на глубине. В зонах с сероводородным заражением осадки обогащены органическим веществом и в ископаемом состоянии дают черносланцевые формации. Надежным критерием установления сероводородного заражения является наличие скоплений планктонных организмов при полном отсутствии бентосной фауны и одновременно повышенное количество органики во вмещающих осадках. Это связано с тем, что в условиях сероводородного заражения не происходит минерализации органического вещества, которое захороняется в осадке.
Движение воды в прибрежной зоне и на морском дне (действие волн, течения, приливы и отливы) заставляет организмы вырабатывать различные приспособления. Это толстые раковины, прочные постройки, способность прикрепляться ко дну путём всверливания или приклеивания. В зонах подводных течений на скалистом грунте обитают прирастающие животные (например, кораллы, строматопораты). Сильное движение воды сказывается на форме их колоний, строении скелета. Появляются плоские и обтекаемые формы. Некоторые кораллы стелются по дну. В ископаемом состоянии можно встретить такое дно древнего моря – поверхность напластования со стелющимися кораллами. Следами сверления, «пеньками» морских лилий, приросшими брюшными створками брахиопод, трубочками спирорбирсов.
Движение воды усиливает газовый обмен, что способствует расцвету жизни. При этом создаётся окислительная среда. В противном случае возникает восста-новительная обстановка. В застойной среде у дна часто развивается сероводородное заражение. Для таких условий характерно обогащение пород органикой и их темно-серый или черный цвет. В окислительных обстановках при свободном доступе кислорода цвет пород обычно желтый или бурый.
Характер грунта во многом определяет способность животных приспосабливаться к жизни и определяет расселение донных животных. Наиболее благоприятен песчаный грунт для перемещающегося, временно или постоянно зарывающегося бентоса. У зарывающихся полностью или частично брахиопод и двустворок изменяются форма раковины, её скульптура и внутреннее строение (рис. 2.3), раковина становится длиннее. Рыхлый грунт менее благопри-ятен, кроме зарывающихся организмов, требует у них дополнительных скелетных элементов. У морских лилий появляются образования, напоминающие корни. Раковины моллюсков и брахиоподы становятся широкими и плоскими, непогружающимися в грунт. Подобной цели служат иглы у некоторых морских ежей, шипы, иглы, выросты на раковинах брахиопод, двустворок и гастропод. В рыхлом грунте сохраняются следы жизнедеятельности илоядных форм, например, ходы червей.
На твёрдом грунте формы при-крепляются с помощью цемента. Многие пелециподы и брахиоподы образуют банки, нарастая друг на друга. Кораллы, археоциаты, строматопораты растут на твёрдом грунте. В него всверливаются разнообразные камнеточцы. Некоторые присасываются к грунту, другие подве-шиваются с помощью биссуса. Обитание на определённом грунте отражается в твёрдых скелетных элементах. Поэтому по остаткам бентосных форм можно судить о типе грунта, на котором они жили.
П ри
биономическом анализе важное значение
имеет изучение характера захоронения
комплекса ископаемых органических
остатков – ориктоценозов. Обычно
различают два типа захоронений:
палеобиоценоз – захоронение на месте
(рис. 2.4) и тафоценоз – результат переноса
(рис. 2.5). В последнем случае органические
остатки связаны общим местом захоронения,
сортированы обычно по крупности.
Скелетные организмы принимают наиболее
устойчивое положение: вытянуты параллельно
движению, раковины ориентированы
выпуклой створкой вверх и т.д. В таком
случае происходит совместное захоронение
остатков животных, обитавших в разных
условиях, например, на скальном и рыхлом
грунтах. При отсутствии переноса остаются
на месте крупные и мелкие экземпляры.
ри
биономическом анализе важное значение
имеет изучение характера захоронения
комплекса ископаемых органических
остатков – ориктоценозов. Обычно
различают два типа захоронений:
палеобиоценоз – захоронение на месте
(рис. 2.4) и тафоценоз – результат переноса
(рис. 2.5). В последнем случае органические
остатки связаны общим местом захоронения,
сортированы обычно по крупности.
Скелетные организмы принимают наиболее
устойчивое положение: вытянуты параллельно
движению, раковины ориентированы
выпуклой створкой вверх и т.д. В таком
случае происходит совместное захоронение
остатков животных, обитавших в разных
условиях, например, на скальном и рыхлом
грунтах. При отсутствии переноса остаются
на месте крупные и мелкие экземпляры.