

5. Регуляция дыхания
Дыхание, наряду с кровообращением, относиться к категории жизненно важных функций организма, которые осуществляются постоянно ритмически. Определяющую роль в периодической смене вдоха выдохом (т.е. в осуществлении дыхательных циклов), а также в регуляции глубины и частоты дыхания играет дыхательный центр. Причем уровень вентиляции легких (глубина и частота внешнего дыхания) во многом зависит от интенсивности обменных процессов в периферических тканях, которая отражается на газовом составе крови. Дыхательный центр постоянно подстраивает интенсивность внешнего дыхания под текущие потребности организма, обеспечивая поддержание газового состава альвеолярного воздуха в таких пределах, при которых достигается нормальное насыщение артериальной крови кислородом. Следовательно, конечным результатом деятельности дыхательного центра является поддержание газового состава артериальной крови на уровне, оптимальном для нормального протекания метаболических процессов во всех тканях организма.
Под дыхательным центром понимают совокупность нервных клеток, заложенных в разных отделах центральной нервной системы и обеспечивающих координированную ритмичную деятельность дыхательных мышц и приспособление дыхания к изменяющимся условиям внешней и внутренней среды организма. Жизненно важный дыхательный центр, который собственно обладает автоматией (способностью к самостоятельной периодической генерации возбуждений) и обеспечивает дыхательную периодику (постоянную смену вдоха выдохом и, как следствие, ритмичное осуществление дыхательных циклов, продолжительность и глубина которых может регулироваться произвольно), заложен в продолговатом мозге (в толще дна ромбовидной ямки, бульбарный дыхательный центр) и представлен двумя парами ядер:
дорсальными дыхательными ядрами (совокупность нейронов, окружающих солитарное ядро), содержат преимущественно инспираторные нейроны, тогда как на долю экспираторных нейронов приходится всего 5% от общего количества нейронов этих ядер. Аксоны инспираторных нейронов вступают в синаптический контакт с мотонейронами инспираторных мышц, которые заложены в шейных (мотонейроны диафрагмы) и грудных (мотонейроны межреберных мышц) сегментах спинного мозга
вентральными дыхательными ядрами (расположены в вентролатеральной области ромбовидного мозга, характеризуются довольно большой протяженностью: от ядра лицевого нерва до I шейного сегмента спинного мозга), включают в свой состав обоюдное ядро (общее двигательное ядро для языкоглоточного и блуждающего нервов, часть нейронов которого имеет отношение к иннервации мышц гортани и глотки). Большая часть нейронов этого ядра имеет отношение к экспирации и посылает свои аксоны к мотонейронам спинного мозга, иннервирующим внутренние межреберные мышцы и мышцы живота.
М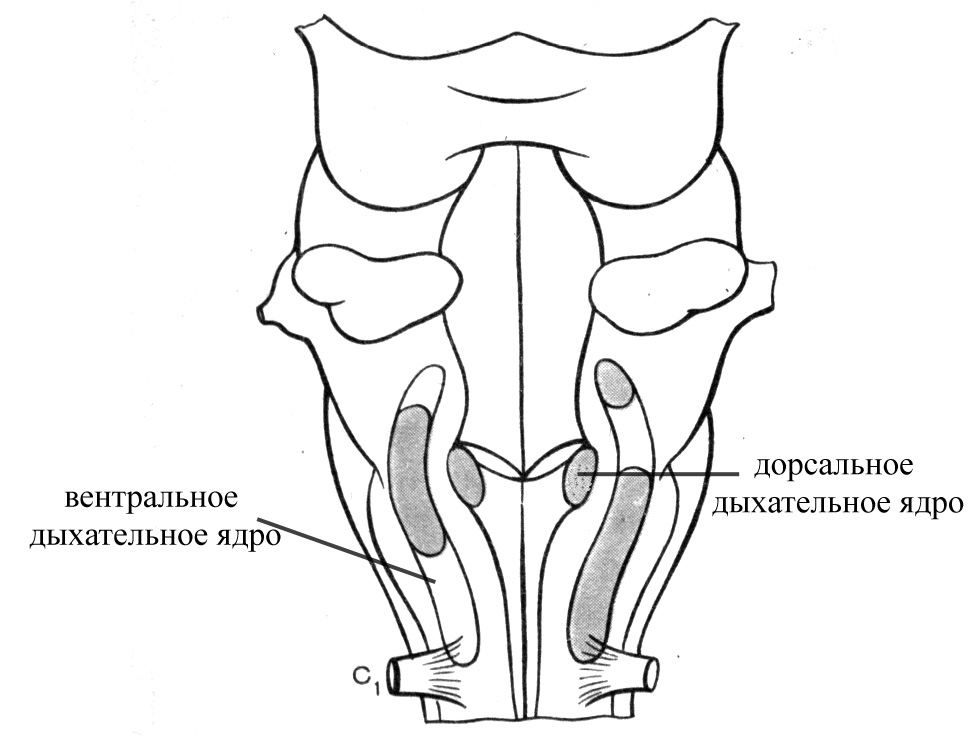 ежду
нейронами этих двух пар дыхательных
ядер существуют сложные взаимоотношения.
Так, инспираторные нейроны дорсального
дыхательного ядра, наряду с тем, что
посылают свои аксоны в спинной мозг (к
мотонейронам инспираторных мышц), отдают
еще и коллатерали, вступающие в
синаптический контакт с нейронами
вентрального дыхательного ядра. Причем
инспираторные нейроны дорсального
дыхательного ядра, возбуждаясь, приводят
в возбуждение и инспираторные нейроны
вентрального дыхательного ядра, но при
этом оказывают тормозное действие на
его экспираторные нейроны. Разрушение
дыхательного центра продолговатого
мозга или нарушение его связи с
мотонейронами дыхательных мышц спинного
мозга приводит к остановке дыхания.
ежду
нейронами этих двух пар дыхательных
ядер существуют сложные взаимоотношения.
Так, инспираторные нейроны дорсального
дыхательного ядра, наряду с тем, что
посылают свои аксоны в спинной мозг (к
мотонейронам инспираторных мышц), отдают
еще и коллатерали, вступающие в
синаптический контакт с нейронами
вентрального дыхательного ядра. Причем
инспираторные нейроны дорсального
дыхательного ядра, возбуждаясь, приводят
в возбуждение и инспираторные нейроны
вентрального дыхательного ядра, но при
этом оказывают тормозное действие на
его экспираторные нейроны. Разрушение
дыхательного центра продолговатого
мозга или нарушение его связи с
мотонейронами дыхательных мышц спинного
мозга приводит к остановке дыхания.
Рис. 11. Ромбовидный мозг, вид сзади (показаны ядра дыхательного центра)
Активность дыхательного центра продолговатого мозга, в свою очередь, находится под регулирующим контролем пневмотаксического центра моста (расположен в передней части моста), который оказывает тормозное влияние на инспираторные нейроны дыхательных ядер продолговатого мозга, тем самым ограничивая (укорачивая) инспирацию. Повреждение пневмотаксического центра моста или перерезка ствола головного мозга между мостом и продолговатым мозгом не приводит к остановке дыхания, но сопровождается значительным удлинением вдохов и некоторым удлинением выдохов (в целом дыхание становится более редким). Таким образом, пневмотаксический центр моста увеличивает скорость развития инспирации, укорачивает ее продолжительность и ускоряет фазу экспирации.
Немаловажное значение в регуляции дыхания имеют и вышерасположенные отделы головного мозга и, прежде всего, гипоталамус, являющийся высшим вегетативным центром, и кора больших полушарий. Так, гипоталамус обеспечивает тонкую подстройку частоты и глубины дыхания текущим потребностям организма при выполнении сложных поведенческих актов. Например, тяжелая физическая нагрузка сопровождается, как правило, усилением дыхания (с целью увеличить его минутный объем), увеличением интенсивности сердечной деятельности, уровня теплоотдачи организмом и некоторыми другими перестройками, возникающими сочетано в результате регулирующего влияния гипоталамуса на определенные ниже расположенные центры этих функций. Корковая же регуляции активности жизненно важного дыхательного центра продолговатого мозга (которая может осуществляться как прямым способом – через кортико-бульбарные пути, так и косвенно – через посредство гипоталамуса, ретикулярной формации и некоторых других структур) обеспечивает возможность
произвольного контроля над дыханием (человек может сам, т.е. произвольно, углублять, учащать ил замедлять дыхание)
условнорефлекторного регулирования дыхания
определенного изменения дыхания при речи и пении, мышлении, эмоциональных состояниях и т.д.
Несмотря на описанное разнообразие структур центральной нервной системы, имеющих отношение к регуляции дыхания, все же определяющую роль в постоянном циклическом его осуществлении играет бульбарный дыхательный центр, который своей деятельностью обеспечивает дыхательную периодику. Активность нейронов бульбарного дыхательного центра, обладающего автоматией, регулируется преимущественно нервным путем.
Нервная регуляция активности дыхательного центра осуществляется рефлекторно в ответ на раздражение различных рецептивных полей организма (механорецепторов аппарата внешнего дыхания, проприорецептров дыхательных мышц, хеморецепторов сосудистого русла, периферических тканей и самого продолговатого мозга, а также некоторых других рецептивных полей). Так, в крупных сосудах (дуге аорты и области каротидного синуса), многих мелких сосудах, самих периферических тканях заложены хеморецепторы (т.н. периферические хеморецепторы), адекватными раздражителями для которых выступают:
уменьшение содержания кислорода в крови (гипоксемия) и тканях (гипоксия)
увеличение содержания углекислого газа в крови (гиперкапния) и тканях
сдвиг рН крови и межклеточной жидкости в кислую сторону.
повышенная концентрация ионов Н+ в крови и межклеточной жидкости.
В самом продолговатом мозге (вблизи дыхательных ядер) находятся т.н. центральные хеморецепторы, возбуждающиеся в ответ на повышение концентрации в церебральной жидкости и артериальной крови углекислого газа и ионов Н+.
Возбуждение как периферических, так и центральных хеморецепторов оказывает активирующее влияние на бульбарный дыхательный центр, способствуя интенсификации дыхания и нормализации газового состава крови. Причем центральные хеморецепторы оказывают более выраженное влияние на деятельность дыхательного центра по сравнению с периферическими.
В стенках воздухоносных путей и некоторых структур респираторного отдела (кроме альвеол) заложены механорецепторы (рецепторы растяжения), адекватным раздражителем для которых является растяжение стенок дыхательных путей. Наличие рецепторов растяжения в аппарате внешнего дыхания обеспечивает возможность объемной обратной его связи с дыхательным центром. Возбуждение механорецептров аппарата внешнего дыхания может вызвать следующие рефлексы (впервые были описаны Герингом и Брейером в 1868г):
инспираторно-тормозящий рефлекс, возникает в ответ на раздувание легких при вдохе и проявляется в прекращении вдоха и возникновении выдоха; имеет защитное значение, поскольку препятствует сильному перерастяжению легких при вдохе
экспираторно-облегчающий рефлекс, возникает в ответ на раздувание легких при выдохе и проявляется в продлении экспирации и задержке очередной инспирации
парадоксальный эффект Хеда, заключающийся в том, что в ответ на сильное возбуждение механорецепторов раздуванием легких кратковременно повышается активность инспираторных мышц и возникает судорожный вдох (вздох), который приводит к расправлению легких и восстановлению равномерности их вентиляции
наконец, снижение активности механорецептров, возникающее во время выдоха, способствует усилению инспираторной активности и укорочению выдоха (рефлекс на спадение легких).
Наряду с типичными механорецепторами (рецепторами растяжения), во всех воздухоносных путях (в эпителиальной выстилке их слизистой оболочки и субэпителиальном слое) залегают ирритантные рецепторы, представляющие собой нечто среднее между механо- и хеморецепторами. Так, адекватными раздражителями для них может выступать как сильное перерастяжение, так и спадение легких, пары едких веществ, пылевые частицы, гистамин и некоторые другие факторы. При раздражении ирритантных рецепторов дыхательных путей пылевыми частицами или едкими веществами возникает кашель. В случае раздражения их спадением бронхов или легких рефлекторно усиливается инспираторная активность, что препятствует дальнейшему спадению легких. Возбуждение этих рецепторов сильным перерастяжением легких, напротив, усиливает экспираторную активность. Возбуждение же этих рецепторов перерастяжением легких в момент пониженной их растяжимости может привести к возникновению парадоксального эффекта Хеда.
Немаловажное значение для регуляции активности дыхательного центра имеют и проприорецепторы самих дыхательных мышц, сигнализирующие в центральную нервную систему о степени их укорочения. Благодаря отмеченной двусторонней связи между центральной нервной системой и дыхательными мышцами, дыхательный центр может постоянно регулировать и корректировать глубину дыхания.
Зависимость деятельности дыхательного центра от газового состава крови
Деятельность дыхательного центра, определяющая частоту и глубину дыхания, находится в прямой зависимости от газового состава крови и тканевой жидкости, а также содержания ионов Н+ в них. Именно газовый состав крови и, прежде всего, парциальное напряжение углекислого газа в ней создает запрос на нужную степень вентиляции альвеол (т.е. определяет интенсивность внешнего дыхания).
Нормальное содержание кислорода в крови носит название нормоксемии, углекислого газа – нормокапнии. Уменьшение парциального напряжения кислорода в крови называется гипоксемией, а в тканях – гипоксией. Увеличение содержания углекислого газа в крови носит название гиперкапнии, а сдвиг рН крови в кислую сторону – ацидоза. Состояние, при котором гипоксия и гиперкапния встречаются одновременно, называется асфиксией.
Гипоксемия, гиперкапния и ацидоз являются адекватными раздражителями хеморецепторов сосудистого русла, а гипоксия и ацидоз тканевой жидкости – тканевых хеморецепторов и первоначально приводят к рефлекторной интенсификации внешнего дыхания (развитию гиперпноэ), направленной на восстановление нормального газового состава крови. Длительная гипоксия или даже кратковременная асфиксия в тканях животного организма и, прежде всего, в головном мозге сопровождается нарушением обменных процессов в нем и может привести к ослаблению дыхания (гипопноэ) или полной его остановке (апноэ). Гипокапния, возникающая обычно вследствие гипервентиляции легких, сопровождается ослаблением дыхания и возможно полной его остановкой по причине снижения активирующего влияния углекислого газа на дыхательный центр. Таким образом, как избыток, так и недостаток углекислого газа (является адекватным раздражителем дыхательного центра) в крови и церебральной жидкости негативно отражается на внешнем дыхании.
Механизм периодической деятельности дыхательного центра
Механизм периодической деятельности дыхательного центра до настоящего времени полностью не раскрыт. Одной из самых общепризнанных моделей его деятельности является следующая. В бульбарном дыхательном центре существует популяция нейронов, которые выступают генераторами центрального инспираторного возбуждения (это I-нейроны или ранние инспираторные нейроны); они возбуждаются сигналами от периферических и центральных хеморецепторов и параллельно подавляют активность экспираторных нейронов. Возбуждение I-нейронов и обуславливает развитие инспирации. Последующее торможение инспирации и возникновение экспирации обеспечивается группой I-нейронов (поздних инспираторных нейронов), возбуждающихся потоком афферентной информации от рецепторов растяжения воздухоносных путей, которые повышают свою активность по ходу вдоха. Группа I-нейронов оказывает тормозное влияние на I-нейроны, которые, в свою очередь, находясь в активном состоянии, подавляли экспираторные нейроны. Следовательно, активация нейронов I афферентной импульсацией, поступающей от механорецепторов аппарата внешнего дыхания, сопровождается торможением нейронов-генераторов центрального инспираторного возбуждения (I-нейронов) и, как следствие, прекращением их тормозного влияния на экспираторные нейроны, что способствует возникновению экспирации. Соответственно основой механизма периодической деятельности дыхательного центра согласно описанной модели считают два процесса:
генерацию центрального инспираторного возбуждения. Обеспечивается возбуждением I-нейронов, возникающим в ответ на афферентную импульсацию, которая поступает от хеморецепторов сосудистого русла и от центральных хеморецепторов. Активные I-нейроны, в свою очередь, оказывают тормозное влияние на экспираторные нейроны
выключение инспирации. Обеспечивается возбуждением I-нейронов, возникающим в ответ на афферентную импульсацию от механорецептров дыхательных путей, подвергающихся растяжению по ходу инспирации. Возбуждение I-нейронов, в свою очередь, подавляет активность I-нейронов (нейронов-генераторов центрального инспираторного возбуждения) и снимает тем самым блок с экспираторных нейронов. Оба эти обстоятельства способствуют экспирации.
возбуждение хеморецепторов сосудистого
русла и центральных хеморецепторов
повышенным содержанием СО2, Н+
и пониженной концентрацией О2

ВОЗБУЖДЕНИЕ