

2.3.Физиологические основы голода и насыщения
Потребность в питательных веществах выражается в состоянии голода и создает мотивацию поиска и поедания пищи. Совокупность нейронов различных отделов центральной нервной системы, которые определяют пищевое поведение и регулируют пищеварительные функции человека и животного, составляют пищевой центр. Между центром голода и центром насыщения существуют реципрокные отношения, т.е. если один центр возбужден, то другой заторможен. Возбуждение или торможение этих ядер происходит в зависимости от содержания питательных веществ в крови, а также сигналов, поступающих от различных рецепторов.
Существует несколько теорий, объясняющих возникновение чувства голода:
Глюкостатическая теория - ощущение голода связано со снижением уровня глюкозы в крови.
Аминоацидостатическая - чувство голода создается понижением содержания в крови аминокислот.
Липостатическая - нейроны пищевого центра возбуждаются недостатком жирных кислот и триглицеридов в крови.
Метаболическая - раздражителем нейронов пищевого центра являются продукты метаболизма цикла Кребса.
Термостатическая - снижение температуры крови вызывает чувство голода.
Локальная теория - чувство голода возникает в результате импульсации от механорецепторов желудка при его "голодных" сокращениях.
Насыщение возникает в результате возбуждения нейронов центра насыщения. Выделяют первичное, или сенсорное, насыщение и вторичное, или обменное. Сенсорное насыщение связано с торможением латеральных ядер гипоталамуса импульсами от рецепторов рта, желудка, возбуждаемых принимаемой пищей. В то же время возбуждение нейронов вентромедиальных ядер гипоталамуса импульсами от рецепторов рта, желудка, возбуждаемых принимаемой пищей.
В то же время возбуждение нейронов вентромедиальных ядер гипоталамуса приводит к поступлению в кровь питательных веществ из депо. Вторичное, обменное, или истинное, насыщение наступает через 1,5-2 часа с момента приема пищи, когда в кровь поступают продукты гидролиза питательных веществ. Гормоны желудочно-кишечного тракта также играют важную роль в возникновении чувства голода и насыщения. Холецистокинин, соматостатин, бомбезин и другие снижают потребление пищи. Пентагастрин, окситоцин и другие способствуют формированию чувства голода.
Контрольные вопросы
1.Что подразумевается под процессом пищеварения?
2. Что называют пищеварительной системой?
3. Перечислите основные функции пищеварительной системы?
4. Назовите основные отделы желудочно- кишечного тракта?
5.Охарактеризуйте суть пищеварительных процессов в ротовой полости?
6. Охарактеризуйте суть пищеварительных процессов в желудке?
7. Охарактеризуйте суть пищеварительных процессов в тонком кишечнике?
8. Охарактеризуйте суть пищеварительных процессов в толстом кишечнике?
9.Охарактеризуйте особенности процессов всасывания основных пищевых веществ в ЖКТ?
10.Назовите механизмы обеспечения регуляции процесса пищеварения?
3. Процессы всасывания и усвоения пищевых веществ
Пищевые вещества содержат крупномолекулярные белки, углеводы и липиды, которые не способны к всасыванию в кровь и лимфу из-за больших размеров своих молекул. Химическая переработка пищи в желудочно-кишечном тракте представляет собой последовательное ступенчатое ферментативное гидролитическое расщепление крупномолекулярных белков, углеводов и липидов до простых веществ, способных к всасыванию. Ферменты, катализирующие эти реакции гидролиза называют гидролазами. Все пищеварительные ферменты синтезируются, резервируются и выделяются в неактивной форме, в виде проферментов и активируются непосредственно перед началом гидролиза.
Переваривание - это гидролитическое расщепление крупных молекул питательных веществ до более мелких, готовых для всасывания - переноса через энтероцит.
При распаде питательных веществ многие их свойства утрачиваются. Благодаря этому, в частности, предотвращается попадание инородного белка в организм.
В транспорте веществ через мембрану энтероцита участвуют 4 механизма:
активный транспорт;
простая диффузия;
облегченная диффузия;
эндоцитоз.
Активный транспорт идет против концентрационного или электрохимического градиента и требует затрат энергии. Этот вид транспорта происходит с участием белка-переносчика; возможно также его конкурентное ингибирование.
Простая диффузия, наоборот, идет по концентрационному или электрохимическому градиенту, не требует затрат энергии, осуществляется без белка-переносчика и не подвержена конкурентному ингибированию.
От простой облегченная диффузия отличается тем, что для нее необходим белок-переносчик и возможно ее конкурентное ингибирование.
Простая и облегченная диффузия - это разновидности пассивного транспорта.
Эндоцитоз напоминает фагоцитоз: питательные вещества, растворенные или в виде частиц, попадают в клетку в составе пузырьков, образованных клеточной мембраной. Эндоцитоз происходит в кишечнике новорожденных, у взрослых он выражен незначительно. Вероятно, именно он обусловливает (по крайней мере, частично) захват антигенов.
Для многих компонентов пищи характерно преимущественное всасывание в определенных ее отделах. В проксимальном отделе всасывается большая часть железа, кальция, жиров (моноглицеридов и жирных кислот) и водорастворимых витаминов. Моно- и дисахариды всасываются в двенадцатиперстной и тощей кишках, аминокислоты - преимущественно в тощей. Желчные кислоты и витамин В12 всасываются в основном в подвздошной кишке, и при ее заболеваниях или резекции этот процесс нарушается. В толстой кишке (преимущественно в слепой) всасываются вода и электролиты. Прямая кишка обычно не участвует во всасывании, но в ней могут всасываться введенные ректально препараты (например, салицилаты или глюкокортикоиды), оказывая как местное, так и общее действие.
Переваривание углеводов
Большая часть углеводов (около 60%) в пище представлена растительным крахмалом, 30% сахарозой, 10% лактозой. В пище содержатся небольшие количества глюкозы и фруктозы, а также гликоген.
Переваривание полисахаридов носит многоступенчатый характер (см. табл. 2). Переваривание крахмала- основного углевода пищевых продуктов – начинается уже в ротовой полости под влиянием амилазы слюны, активной в условиях нейтрального или щелочного рН слюны. Однако кратковременность пребывания пищи в ротовой полости и относительно низкая активность амилазы слюны делают этот этап переваривания крахмала малоэффективным. Хотя надо заметить содержание этого фермента в слюне очень значительно.
Таблица 2
Переваривание углеводов - основные процессы
Субстрат и конечные продукты |
Фермент и место его выработки |
Механизм действия |
Крахмал до олигосахаридов и амилопектин |
Слюнные железы альфа-амилаза |
Расщепляет альфа-1,4-связи амилозы в составе крахмала опт. рН 6,7 |
Крахмал до олигосахаридов |
Поджелудочная железа Панкреатическая амилаза |
Расщепляет альфа-1,4-связи амилозы в составе крахмала опт. рН 7,1 |
Крахмал и олигосахариды до мальтозы и глюкозы |
Ферменты, связанные с мембраной энтероцитов амилаза |
Глюкоамилаза |
Гликоген, амилопектин до олигосахаридов, мальтозы, глюкозы |
олиго-альфа1,6-глюкозидаза |
Расщепляет альфа-1,6связи амилопектина |
Сахароза до фруктозы и глюкозы |
Дисахаридазы Сахараза |
Бета-фруктозидаза |
Мальтоза до глюкозы |
Мальтаза |
Альфа-глюкозидаза, расщепляет альфа-1,4-связи |
Мальтоза до глюкозы |
Изомальтаза |
Действует аналогично альфа-1,6-глюкозидазе |
Лактоза до галактозы и глюкозы |
Лактаза |
Бета-галактозидаза |
В желудке амилаза инактивируется кислым содержимым желудка, и переваривание углеводов прекращается. И только в 12-перстной кишке происходит полный гидролиз крахмала, включая и образовавшийся в полости рта альфа- лимит-декстрин, и всех дисахаридов до моносахаров. Гидролиз углеводов в кишечнике осуществляется ферментами поджелудочной железы (альфа- амилаза, олиго – 1,6 - глюкозидаза) и кишечника (олигосахаридазы, дисахаридазы).
Эффективность расщепления крахмала под влиянием амилазы и глюкоамилазы зависит от ряда факторов, касающихся как особенностей формы крахмала в пищевых продуктах, так и функционального состояния ЖКТ.
В последние годы, было установлено, что существует так называемые резистентные формы крахмала, устойчивые к ферментативному расщеплению в кишечнике: они расщепляются значительно медленнее. Существование таких резистентных форм крахмала обусловлено двумя основными причинами:
Способностью крахмала образовывать достаточно прочные комплексы с растительными волокнами, белками, другими компонентами клеток и клеточными структурами с формированием при этом физически защищенных форм крахмала и крахмальных гранул, в которых крахмал трудно доступен для расщепления ферментами ЖКТ человека.
Нестабильностью желатинизированной формы крахмала, возникающей при его нагревании в присутствии воды. Образование этой формы крахмала сопровождается разрушением крахмальных гранул и быстрым ферментативным расщеплением крахмала. Эта нестабильность процесса желатинизации приводит к тому, что при охлаждении продукта, предварительно подвергнутого термической обработке (варка картофеля, выпечка хлеба), либо при некоторых видах технологической обработки зерновых происходит процесс, обратный желатинизации, и вновь образуются крахмальные гранулы, в которых крахмал оказывается трудно доступным для ферментной атаки. Важно подчеркнуть, что амилоза в большей степени способна к реассоциации в крахмальные гранулы. Поэтому продукты, содержащие большие количества амилозы, хуже расщепляются амилазой, о чем свидетельствует различия в гликемических индексах таких продуктов.
Дисахариды предварительно расщепляются до моносахаридов под влиянием соответствующих дисахаридаз – сахаразы, лактазы и мальтазы, секретируемых в кишечнике, и всасываются в основном в виде моносахаридов. Гидролиз лактозы идет медленнее, и поэтому именно он ограничивает скорость ее всасывания.
Дисахариды гидролизуются не в полости, а в стенке кишечника, поэтому образующиеся моносахариды сразу всасываются.
Всасывание моносахаридов галактозы и глюкозы происходит в два этапа с помощью активного транспорта. Прежде всего сахаридазы, расположенные в щеточной каемке энтероцитов, расщепляют олигосахариды до моносахаридов, которые переносятся в клетку с участием системы натрий - зависимого транспорта. При этом моносахариды в присутствии ионов натрия связываются с переносчиком. Присоединив натрий и глюкозу, этот переносчик диффундирует по электрохимическому градиенту для ионов натрия к внутренней стороне мембраны. Затем он высвобождает ион натрия и глюкозу в цитоплазму и диффундирует обратно к наружной поверхности энтероцита. Сравнительно низкое содержание натрия в клетке поддерживается благодаря действию энергозависимого натриевого насоса, работа которого косвенно способствует постоянной диффузии переносчика, связанного с натрием, к внутренней стороне мембраны.
Манноза и пентозы поступают в клетку путем простой, а фруктоза - путем облегченной диффузии (пассивный транспорт).
Высвобождение моносахаридов в области боковой и базальной поверхности энтероцита, по современным представлениям, не зависит от ионов натрия.
Выделившиеся моносахариды удаляются от кишечника по ветвям воротной вены.
Главные потребителями глюкозы помимо печени являются головной мозг, скелетные мышцы. В жировой ткани глюкоза используется для синтеза жировой ткани. Обычно около 65% глюкозы, поступившей при всасывании из кишечника, расходуется на окисление в клетках, на синтез жира около 30% и 5% на синтез гликогена. Эти пропорции меняются в зависимости от физиологического состояния организма, возраста и ряда других причин.
Переваривание и всасывание белков
Переваривание белков в желудочно-кишечном тракте - это часть метаболизма белков, совокупность управляемых процессов, осуществляемых в системе пищеварения, которые представляют собой химическую переработку белков поступающих в организм с пищей, для последующего их всасывания в кровь и в лимфу. Переваривание белков пищи предназначено для их денатурации, лишения видовой и тканевой специфичности и расщепления на простые компоненты способные всасываться в тонкой кишке в кровь. Еще в 50-х годах 20 века было установлено, что протеолизу подвергается только денатурированный белок. О степени перевариваемости белка судят по его перевариваемости протеазами. Нативный белок не подвергается протеолизу. В противном случае функционирование активных белков и само их существование было бы невозможным, поскольку протеолитические ферменты как участники белкового обмена присутствуют в каждой клетке. В живых организмах протеазы играют роль санитаров, расщепляя неактивные, денатурированные молекулы.
Почти все белки пищи (~95÷97%) всасываются в виде свободных аминокислот.
Расщепление белков пищи представляет собой гидролиз с участием катализаторов - протеолитических ферментов (протеазы, протеиназы, пептидазы). Каждый фермент из протеиназ разрывает вполне определенные пептидные связи белков. Специфичность действия зависит от размера полипептида, его структуры, разновидности аминокислот участвующих в образовании пептидных связей.
В зависимости от особенностей действия, пептидазы разделяют на две группы экзопептидазы и эндопептидазы. Экзопептидазы катализируют разрыв концевой пептидной связи. При этом высвобождается вполне определенная концевая аминокислота.
Эндопептидазы гидролизуют главным образом пептидные связи внутри полипептидной цепи. Разные эндопептидазы обладают специфичностью действия на субстрат гидролиза, зависящей от вида аминокислот в разрываемой пептидной связи. В связи с этим под действием разных эндопептидаз молекула белка расщепляется на определенное количество пептидов.
Переваривание белков пищи начинается в желудке (см. рис.8). Желудок является резервуаром, где потребленная пища находится в течение ~6÷8ч. Ёмкость желудка взрослого человека в среднем равна ~3л (1,5÷4,0л). В желудок поступает пища, которая подверглась существенной физической и незначительной химической переработке в полости рта. Химическая переработка пищи более интенсивно продолжается в желудке. Здесь происходит её постепенное перемешивание с желудочным соком и образование химуса. Желудочный сок содержит соляную кислоту и ферменты, катализирующие гидролиз пищевых веществ, главным образом белков. Слои химуса, прилегающие к стенке желудка, подготовленные в результате начального гидролиза для дальнейшего еще более интенсивного переваривания, периодически порциями перемещаются желудком в двенадцатиперстную кишку.
Соляная кислота желудочного сока играет существенную роль в переваривании белков. Она способствует набуханию белков, их денатурации, создает оптимальную кислотность среды для наилучшего действия ферментов, активирует проферменты, стимулирует секрецию гормонов принимающих участие в управлении функциями желудочно-кишечного тракта.
Желудочный сок содержит три главных протеазы: пепсин, реннин и гастриксин. Пепсин катализирует гидролиз пептидных связей почти всех натуральных белков (за исключением некоторых кератинов, протаминов, гистонов и мукопротеинов). В результате гидролиза белков образуются различного размера полипептиды, олигопептиды и небольшое количество свободных аминокислот.
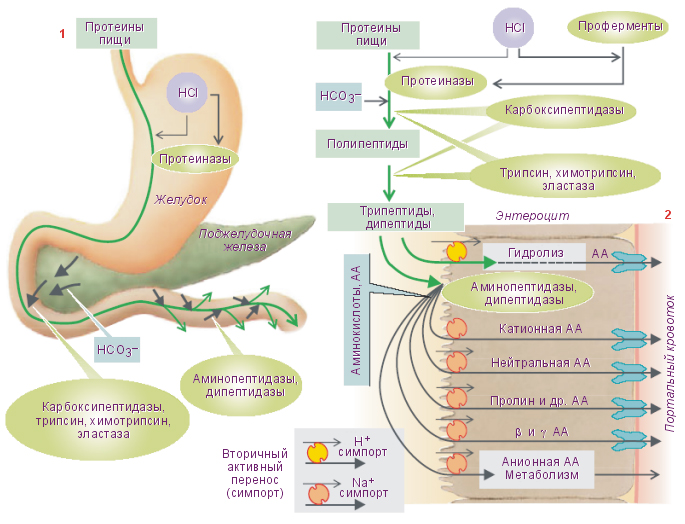
Рисунок 8. Переваривание белков пищи и всасывание аминокислот и олигопептидов в желудочно-кишечном тракте
В желудочном соке детей грудного возраста содержится фермент реннин. Он катализирует свертывание молока (превращение растворимого казеиногена в нерастворимый казеин). При таком свертывании молока замедляется продвижение нерастворимого казеина по желудочно-кишечному тракту. Благодаря этому увеличивается время действия протеиназ и обеспечивается эффективность усвоения белков. У взрослых свертывание молока осуществляется в большей степени пепсином.
Значительно более интенсивное, чем в желудке переваривание белков осуществляется в тонкой кишке. Начальным отделом её является двенадцатиперстная кишка. Двенадцатиперстная кишка имеет приблизительно в десять раз меньшую ёмкость, чем желудок. В то же время через двенадцатиперстную кишку проходит вся та масса химуса, которая находится в желудке. В полость двенадцатиперстной кишки выделяют свои секреты поджелудочная железа, бруннеровы железы и печень. Количество секретов на единицу поверхности двенадцатиперстной кишки, количество ферментов значительно больше, чем на единицу поверхности желудка. Все это объясняет тот факт, что интенсивность переваривания пищевых продуктов, в том числе и белков в двенадцатиперстной кишке приблизительно в четыре раза больше, чем в желудке.
Сок поджелудочной железы содержит пять главных ферментов: трипсин, химотрипсин, коллагеназу, карбоксипептидазу, эластазу. Трипсин и химотрипсин действуют на белки аналогично пепсину. Они наиболее активны в слабощелочной среде (рН = 7,2÷7,8) и разрушают внутренние пептидные связи. Таким образом, внутренние пептидные связи сложных белков последовательно разрывают три протеазы: пепсин желудочного сока, трипсин и химотрипсин сока поджелудочной железы. В результате образуются полипептиды различной длины и небольшое количество свободных аминокислот.
Дальнейший гидролиз полипептидов до свободных аминокислот осуществляется под влиянием группы более специфичных ферментов - пептидаз. Это карбоксипептидазы (карбоксиполи‑, карбокситри‑, карбоксидипептидазы) и аминопептидазы (аминополи‑, аминотри‑, аминодипептидазы). Эти ферменты разрушают концевые пептидные связи с образованием олигопептидов или свободных аминокислот (в соответствии с названием).
Кроме перечисленных ферментов сок поджелудочной железы содержит ферменты эластазу и коллагеназу. Они катализируют гидролиз эластина и коллагена. Как правило, конечные стадии гидролиза пищевых продуктов осуществляются в гликокаликсе и на мембране энтероцита (мембранное переваривание).
Образовавшиеся при этом простые вещества, в частности свободные аминокислоты, здесь же всасываются через мембрану энтероцита в его цитозоль, затем - в интерстициальное пространство микроворсинок, и далее - в кровь их микрогемациркуляторного русла. Большая часть аминокислот, всосавшихся в микрогемациркуляторное русло кишечных ворсинок, попадает с потоком крови через воротную вену в печень. Небольшое их количество поступает по лимфатическим сосудам в венозную систему. Аминокислоты всасываются посредством механизмов активного трансмембранного транспорта.
Нейтральные (не имеющие электрического заряда) аминокислоты и анионные («кислые», сопряженные кислоты) L–аминокислоты транспортируются по механизму вторичного (сопряженного) активного однонаправленного транспорта (симпорт) вместе с ионами натрия Na+. Так из полости тонкой кишки (в меньшей степени) и с поверхности гликокаликса (в большей степени) аминокислоты попадают в цитозоль энтероцитов. Отсюда они посредством пассивной диффузии или с помощью переносчиков транспортируются в кровь микрогемациркуляторного русла микроворсинок.
Катионные («основные», сопряженные основания) L–аминокислоты, такие как L–аргинин+, L–лизин+, L–орнитин+ могут частично перемещаться в цитоплазму энтероцита посредством механизмов, независимых от транспорта натрия. В этом случае движущей силой для их перемещения является электрический потенциал мембраны энтероцита.
Анионные аминокислоты, такие как L–глютамат– и L–аспартат имеют собственные механизмы транспорта, сопряженные с транспортом натрия Na+ и калия K+. По большей части эти аминокислоты в цитоплазме эритроцита разрушаются. Дипептиды и трипептиды могут транспортироваться в цитоплазму энтероцита посредством механизмов вторичного активного трансмембранного транспорта. Скорость такого транспорта значительно меньше, чем скорость транспорта свободных аминокислот. В цитоплазме энтероцитов дипептиды и трипептиды гидролизуются с образованием свободных аминокислот.
Всасывание продуктов гидролиза полинуклеотидов происходит путем пассивного или облегченного транспорта. Наряду с азотистыми основаниями через мембраны хорошо проникают и нуклеозиды. Поэтому в виде нуклеозидов всасывается часть продуктов переваривания нуклеиновых кислот.
Переваривание жиров
Человек потребляет в день около 60-100 г жиров. Всасываемость и усвояемость жиров зависит от состава жирных кислот и температуры их плавления.
В зависимости от температуры плавления жиры можно разделить на три группы по степени усвояемости:
1)жиры, температура плавления которых ниже 370С, а усвояемость составляет 97—98 %. К ним относят все жидкие растительные жиры, жиры молока, свиное, топленое и гусиное сало, жиры птиц и различных рыб;
2)жиры, температура плавления которых равна 37—500С, а усвояемость составляет около 90 %. К ним относится тканевый жир рогатого скота;
3)жиры, температура плавления которых равна 50—600С, а усваиваются они плохо. К ним относится бараний и говяжий жир.
Примерно 89-90% жиров пищи – это триглицериды, большую часть которых составляют липиды, содержащие жирные кислоты с длинной цепью (16,18 атомов углерода). Очень небольшую часть составляют триглицериды с короткой (2-4 ат. углерода) и средней цепью (6-8 ат. углерода). Остальные 9-10% жиров пищи приходится на фосфолипиды, эфиры холестерола и жирорастворимые витамины.
Переваривание липидов происходит в тех отделах ЖКТ, где имеются обязательные условия:
наличие липолитических ферментов, гидролизующих липиды;
условия для эмульгирования липидов;
оптимальный рН (нейтральная или слабощелочная) среды для действия липолитических ферментов.
В желудке жир дробится на капельки размером около 100 нм. У взрослого человека сильнокислая среда инактивирует желудочную липазу. В кишечнике нейтрализуется поступающая из желудка пища, и жир подвергается эмульгированию. А именно, попадая в двенадцатиперстную кишку, жир и соляная кислота вызывают выброс соответственно холецистокинина и секретина, стимулирующих выделение желчи и панкреатического сока. Компоненты этих двух секретов - желчные кислоты с одной стороны, липаза и колипаза панкреатического сока с другой - обеспечивают переваривание и всасывание жиров.
Желчные кислоты образуются в печени из холестерина в количестве 0,2-0,6 г/сутки и попадают в желчь в конъюгированном виде (с глицином и таурином). В основном образуются конъюгаты холевой кислоты и хенодезоксихолевой кислоты. В подвздошной кишке всасывается до 90% поступивших туда конъюгированных желчных кислот. Затем они попадают в воротную вену и возвращаются в печень: происходит кишечно-печеночный кругооборот. За сутки весь запас желчных кислот (3-4 г) проходит через кишечник 5-10 раз (то есть в двенадцатиперстную кишку в сутки поступает 20-30 г желчных кислот), но лишь 0,2-0,6 г выводится с калом.
При заболеваниях или резекции подвздошной кишки всасывание желчных кислот нарушается и возрастает их потеря с калом. В итоге их концентрация в кишечнике падает, что приводит к нарушению всасывания жира.
Желчные кислоты обладают высокой поверхностной активностью. Неполярные (гидрофобные) группы их молекул присоединяются к жирам, и в результате капли жира становятся окруженными слоем желчных кислот, полярные (гидрофильные) группы которых обращены наружу. Благодаря этому на молекулы жиров, расположенные на поверхности этих капель, может действовать гидрофильная липаза. Кроме того, желчные кислоты очищают поверхность капли жира от экзогенных и эндогенных белков.
Колипаза (белок панкреатического сока, присутствующий в нем в виде проколипазы) удерживает липазу у поверхности капли. Без колипазы липаза «смывалась» бы желчными кислотами. Липаза, колипаза и желчные кислоты вместе образуют комплекс, гидролизующий жир. Основные конечные продукты гидролиза - 2-моноглицериды и жирные кислоты, менее 5% жира остается в виде ди- и триглицеридов. При той концентрации желчных кислот, которая создается в кишечнике на высоте пищеварения (5-15 ммоль/л), они соединяются в так называемые мицеллы. В них проникают жирные кислоты и моноглицериды, образуя смешанные мицеллы. Это способствует удержанию жирных кислот и моноглицеридов в растворе (именно поэтому взвесь триглицеридов мутная, а смешанных мицелл - прозрачная). Образование мицелл лучше всего идет при участии конъюгированных желчных кислот и при нормальном рН кишечного содержимого.
В составе смешанных мицелл моноглицериды и жирные кислоты свободно проходят через неподвижный слой жидкости, покрывающей энтероцит, а затем диффундируют в клетку, покидая мицеллу.
В двенадцатиперстной кишке одновременно существуют крупные смешанные мицеллы, насыщенные продуктами липолиза, и еще более крупные жидкокристаллические липосомы, насыщенные свободными жирными кислотами и желчными кислотами. Эти состояния могут переходить друг в друга. Попав в энтероцит, жирные кислоты связываются с особыми белками, и дальнейшая их судьба зависит от длины цепи.
Длинноцепочечные жирные кислоты (16 и 18 ат. углерода) и содержащие их моноглицериды сразу этерифицируются в триглицериды ферментами эндоплазматического ретикулума. Далее вместе с холестерином, фосфолипидами и апопротеинами они образуют хиломикроны и ЛПОНП, которые накапливаются в аппарате Гольджи и секретируются в лимфатические капилляры.
До 30% триглицеридов содержащих жирные кислоты с короткой и средней длиной цепи углеродных атомов, захватываются клетками в интактном виде. Внутри клетки жирные кислоты отщепляются под действием эстераз и вместе с жирными кислотами, поступившими в энтероциты в свободном виде, диффундируют из клеток и поступают через капилляры в воротную вену. Этерифицируется и участвует в образовании липопротеидов лишь небольшая их часть.
В тонком кишечнике происходит образование и так называемых эндогенных триглицеридов (то есть тех, которые синтезируются из эндогенных жирных кислот), однако их главным источником является печень, откуда они секретируются в форме липопротеинов очень низкой плотности (ЛПОНП). В норме всасывается свыше 90% триглицеридов. Это означает, что ежедневно в кровь попадает около 70-150 г экзогенных триглицеридов.
Спектр остатков жирных кислот, обнаруживаемых в триглицеридах хиломикрон и ЛПОНП, в значительной степени зависит от набора жирных кислот триглицеридов, поступающих с пищей. Если в ней, к примеру, оказывается недостаточное количество линолевой кислоты, то в организме может возникать ее дефицит, особенно у пациентов, страдающих нарушениями всасывания. Время полужизни триглицеридов в плазме относительно невелико - они быстро гидролизуются и захватываются различными органами, главным образом, жировой тканью. Эти процессы протекают с участием липолитических ферментов. После приема жирной пищи уровень триглицеридов заметно повышается и остается высоким в течение нескольких часов. В норме все триглицериды хиломикрон должны быть удалены из кровотока в течение 12 часов.
Одновременно с распадом триглицеридов происходит гидролиз холестеридов до холестерина и свободных жирных кислот под действием холестеразы, для которой оптимум рН 6,6 - 8. Холестераза действует главным образом на ненасыщенные жирные кислоты.
В составе хиломикронов и ЛПОНП холестерин попадает в лимфу. Независимо от того, сколько холестерина попадает в организм с пищей, усваивается в среднем 35-40%, причем процесс всасывания опосредуется лимфатической системой. Всасывание холестерина пищи и реабсорбция желчных кислот играют важную роль в ограничении скорости синтеза холестерина клетками печени.
Панкреатическая липаза гидролизует эфиры холестерина, содержащиеся в пище и желчи. Гидролиз завершает холестеринэстераза микроворсинок, всасывается только свободный холестерин. В энтероците большая часть его этерифицируется. Кроме того, энтероциты синтезируют часть эндогенного холестерина.
Фосфолипиды (преимущественно лецитин) расщепляется фосфолипазами А и В. Фосфолипаза А выделяется поджелудочной железой в виде зимогена и в дальнейшем активируется трипсином. Она специфически действует на эфирные связи (в положении 2) лецитина, вызывая его гидролитическое расщепление на лизолецитин и жирные кислоты.
Всасывание витамина A, витамина D, витамина Е и витамина К изучено неполно.
После переноса в энтероцит (или образования в энтероците из бета-каротина) витамин A соединяется в основном с пальмитиновой кислотой, в составе хиломикронов попадает в лимфу и запасается в печени в виде пальмитата.
Витамин D , витамин Е и К также попадают в хиломикроны, но для их транспорта этерификация, очевидно, не требуется.
В различных отделах кишечника вода и электролиты всасываются по-разному. Они могут проходить как через энтероциты (пересекая две мембраны - апикальную и базолатеральную), так и между ними, в обоих случаях попадая в межклеточное пространство. Апикальные отделы соседних клеток связаны плотными контактами, между которыми находятся поры. Обычно закрытые, поры расширяются при всасывании. Апикальная мембрана энтероцитов, образующая микроворсинки, содержит белки-переносчики.
Вода и минеральные соли.
Вода и соли всасываются главным образом в верхних отделах тонкого кишечника. Здесь всасывается большая часть воды, поступившей при питье и в составе пищевых продуктов, а также выделившейся с пищеварительными соками.
В среднем за сутки через тонкий кишечник проходит около 9 л жидкости. Приблизительно 2 л поступает из крови, 7 л с эндогенными секретами желез и слизистой кишечника. Более 80% этой жидкости всасывается обратно в тонком кишечнике – около 60% в двенадцатиперстной кишке и 20% в подвздошной кишке. Остальная жидкость всасывается в толстом кишечнике и только 1% выделяется из кишечника с каловыми массами.
Когда секреция воды и электролитов в тонкой или толстой кишке превышает их всасывание, возникает понос. Вода может диффундировать по обе стороны кишечной стенки как в тонком кишечнике и толстом кишечнике, так (в меньшей степени) и в желудке. Поэтому содержимое кишечника изотонично по отношению к плазме крови. В том случае, когда химус быстро поступает в двенадцатиперстную кишку, ее содержимое может временно становиться гипертоничным, что приводит к всасыванию воды в двенадцатиперсную кишку. Напротив, когда в процессе пищеварения осмотически активные вещества всасываются из кишечника, вода следует за ними по градиенту осмотического давления.
Всасывание Na+ одна из чрезвычайно важных функций тонкого кишечника. Именно за счет ионов Na+ создается в основном электрический и осмотический градиенты; кроме того, они участвуют в сопряженном транспорте других веществ. Всасывание Na+ в кишечнике происходит как за счет активного, так и за счет пассивного механизмов, в том числе путем электрогенного транспорта, сопряженного с переносом незаряженных соединений, электронейтрального обмена и конвекции.
При электрогенном транспорте ионы Na+ переносятся через базолатеральную область мембраны в межклеточное пространство с помощью натриевого насоса, получающего энергию за счет гидролиза АТФ. Это главный механизм всасывания ионов Na+ в кишечнике.
При сопряженном транспорте ионов Na+ незаряженные вещества (D –гексозы, L- аминокислоты, водорастворимые витамины) переносятся в клетку вместе с ионами Na+ общими переносчиками. Таким образом, активный транспорт Na+ косвенным путем обеспечивает энергией процесс, всасывания органических веществ.
При электронейтральном транспорте NaCl в клетку одновременно переносятся ионы Na+ и Сl-, в результате чего процесс и является электронейтральным.
Исключительно важную роль во всасывании ионов Na+ в тонком кишечнике играет пассивный транспорт путем конвекции. Благодаря довольно значительной проницаемости эпителия до 85% ионов Na+ поглощается по механизму «следования за растворителем». При определенной концетрации глюкозы ее всасывание создает ток воды, с которым ионы Na+ и переносятся через межклеточное пространство.
Ионы К+ в отличие Na+ всасываются преимущественно за счет пассивного транспорта по градиенту концентраций. Ионы Сl- всасываются частично вместе с ионами Na+, этому процессу способствует трансэпителиальный электрический градиент. Около 40% ионов Са2+ всасываются в верхнем отделе тонкого кишечника. При низких концентрациях Са2+ всасывание происходит путем активного транспорта, а при высоких концетрациях включается механизм пассивного транспорта. Механизмы всасывания Mg2+ аналогичны всасыванию кальция . Mg2+ подавляет всасывание кальция по типу конкурентного ингибирования, что, возможно, свидетельствует о наличии общей системы переноса этих ионов.
Баланс железа в организме целиком зависит от его всасывания в кишечнике, т.к. специального механизма регуляции его выведения не существует. Железо, поступившее с пищей, всасывается преимущественно в двухвалентном виде. В пищевых продуктах содержатся восстанавливающие вещества, которые могут превращать трехвалентное железо в двухвалентное.
Железо всасывается в верхних отделах тонкого кишечника путем активного транспорта. В энтероцитах железо соединяется с белком апоферритином, образуя ферритин, который служит основным депо железа в организме.
Железо может всасываться, только когда оно находится в виде растворимых комплексов. В кислой среде желудка образуются комплексы железа с аскорбиновой кислотой, желчными кислотами, аминокислотами, моно- и дисахаридами; они остаются в растворенном виде и при более высоком рН двенадцатиперстной и тощей кишок.
В сутки с пищей поступает 15-25 мг железа, а всасывается лишь 0,5-1 мг у мужчин, 1-2 мг у женщин детородного возраста.
Витамин В1 и витамин В2, видимо, всасываются путем простой диффузии.