

Эпиморфоз – процесс регенерации от раневой поверхности – достраивание части до целого. Его смысл заключается в отрастании нового органа от ампутационной поверхности. Выделяют регрессивную и прогрессивную стадии. Регрессивная – заживление раны, разрушение кости и мягких тканей, миграция регенерирующих нервных волокон. Затем начинается прогрессивная стадия – рост и морфогенез. Пример, восстановление конечностей тритона. При эпиморфозе бывает атипичная регенерация – неточная копия. Она бывает:
Гипоморфоз – регенерация с частичным замещением. Пример, у взрослой шпорцевой лягушки возникла шиповидная структура вместо конечности.
Гетероморфоз – появление иной структуры на месте утраченной. Пример, появление конечности на месте антенн или глаза у членистоногих
Избыточная – образуется дополнительная структура. Пример, после надреза культи при ампутации головного отдела планарии возникает регенерация двух голов и более.
Морфаллаксис – оставшаяся после ампутации часть органа перестраивается, превращаясь в целый орган, но меньших размеров. Перестройка органа до целого из остатков. Пример, регенерация конечностей таракана, регенерация гидры из кольца, вырезанного из середины ее тела.
Регенерационная гипертрофия - для восстановления внутренних органов. Этот способ регенерации заключается в увеличении размеров остатка органа без восстановления исходной формы. Раневая поверхность заживает рубцом, удаленный участок не отрастает. И форма органа не восстанавливается. Но увеличивается количество клеток и их размеры, масса органа и его функции восстанавливаются. Пример, регенерация печени, легких, почки.
Компенсаторная гипертрофия – изменение в одном из органов при нарушении в другом, относящемся к той же системе органов. Пример, гипертрофия в одной из почек при удалении другой.
Регуляция репаративной регенерации:
Концепция регенерационных полей – нервная регуляция – при отведении нерва появляется соответствующая часть. Тип нерва роли не играет. Пример, если нерв тритона к основанию хвоста, то образуется дополнительный хвост.
Гуморальная регуляция – после введения нормальным животным плазмы крови от животных, которым удалили печень, у первых наблюдалась стимуляция митотической активности клеток печени
Иммунологический ответ. Не только частичное удаление органа, но и многие воздействия вызывают возмущение в иммунном статусе организма, появление аутоантител и стимуляцию процессов клеточной пролиферации.
46. Особенности регенераторных процессов у млекопитающих и человека. Клеточные источники регенерации. Регенерационная терапия.
Регенерация у млекопитающих отличается своеобразием. Для регенерации некоторых наружных органов нужны особые условия. Язык, ухо, например, не регенерируют при краевом повреждении. Если же нанести сквозной дефект через всю толщу органа, восстановление идет хорошо. В некоторых случаях наблюдали регенерацию сосков даже при ампутации их по основанию. Регенерация внутренних органов может идти очень активно. Из небольшого фрагмента яичника восстанавливается целый орган. Об особенностях регенерации печени уже было сказано выше. Различные ткани млекопитающих тоже хорошо регенерируют. Есть предположение, что невозможность регенерации конечностей и других наружных органов у млекопитающих носит приспособительный характер и обусловлена отбором, поскольку при активном образе жизни нежные морфогенетические процессы затрудняли бы существование. Достижения биологии в области регенерации успешно применяются в медицине. Однако в проблеме регенерации очень много нерешенных вопросов.
Регенерация у человека. У человека хорошо регенерирует эпидермис, к регенерации способны также такие его производные, как волосы и ногти. Способностью к регенерации обладает также костная ткань (кости срастаются после переломов). С утратой части печени (до 75 %), щитовидной или поджелудочной железы клетки оставшихся фрагментов начинают усиленно делиться и восстанавливают первоначальные размеры органа. Нервные клетки также обладают такой способностью. При определённых условиях могут регенерировать кончики пальцев. В связи с обнаружением на регенерирующих тканях слабых электрических напряжений можно предположить, что слабые электрофорезные токи ускоряют регенерацию.
Большие разногласия существуют по вопросу о клеточных источниках регенерации. Существует три предположения.
1. Гипотеза резервных клеток подразумевает, что предшественниками регенерационной бластемы являются так называемые резервные клетки, которые останавливаются на некоем раннем этапе своей дифференцировки и не участвуют в процессе развития до получения стимула к регенерации.
2. Гипотеза временной дедифференцировки - из высоко дифференцированных превращаются в недифференцированные, а затем снова дифференцируются.
3. Гипотеза полной дедифференцировки специализированных клеток до состояния, сходного с мезенхимными клетками и с возможной последующей трансдифференцировкой или метаплазией, т.е. превращением в клетки другого типа, полагает, что в этом случае клетка утрачивает не только специализацию, но и детерминацию.
Регенеративная медицина Говорят, нервные клетки не восстанавливаются, это уже не совсем верно. Стволовые клетки можно превратить в любые клетки человеческого организма, например, заменить шрам на здоровую ткань. В течение всей жизни у человека имеется небольшое число собственных стволовых клеток. В процессе взросления человека наблюдается катастрофическое снижение их количества.
Стволовые клетки способны сами находить сбои в работе нервной, эндокринной, гормональной и т.д. систем и устремляться именно туда и восполнять собою утраченные или поврежденные клетки. Но теперь возможно не только искусственно вводить дополнительные стволовые клетки (не факт, что они при этом сами начнут работать), но и есть попытки "программировать" т.е. задавать им заранее заданную специализацию, направленность. Таким образом, можно достигнуть не просто процесса общего оздоровления, а усиленно воздействовать на ту или иную поврежденную систему организма.
В случае болезни или ранения стволовые клетки могут быть использованы для восстановления или замещения любых поврежденных тканей. Работы по изучению начаты сравнительно недавно, но темпы открытий в этой области чрезвычайно высоки, многие полагают, это будущее медицины.
Уже сегодня революционные технологии, изменяют подходы к лечению многих тяжелых заболеваний.
47. Гомеостаз. Механизмы поддержания гомеостаза: регулирующая система, регуляция по возмущению, регуляция по отклонению, виды обратной связи, примеры.
Гомеостаз – это относительное постоянство внутренней среды организмов и других биосистем в процессе их функционирования и при наличии внешних или внутренних возмущений.
На каждом уровне используются свои механизмы поддержания гомеостаза.
Механизмы поддержания гомеостаза
Организм является самоорганизующейся системой. Основой физиологической регуляции является передача и переработка информации. Информация - это все, что несет в себе отражение фактов или событий, которые произошли, происходят или могут произойти. Переработка информации осуществляется управляющей системой – системой регуляции. Она состоит из отдельных элементов, связанный информационными каналами. Среди элементов выделяются:
1) датчики, воспринимающие информацию на входе системы (сенсорные рецепторы)
2) входные и выходные каналы связи (нервы, жидкости внутренней среды)
3) управляющее устройство (ЦНС)
5) образования, располагающиеся на исполнительных органах.
Регуляция по отклонению (саморегуляция по выходу системы) – обеспечивается сравнением имеющихся физиологических параметров с требующимися в конкретной ситуации, определением рассогласования между ними и включением исполнительных устройств для устранения этого рассогласования.
Регуляция по отклонению требует наличия канала связи между выходом системы регуляции и ее центральным аппаратом управления. Этот канал получил названия обратной связи. По конечному эффекты регуляции обратная связь может быть положительной и отрицательной.
Положительная обратная связь – выходной сигнал системы регуляции усиливает входной. Такая обратная связь усиливает процессы жизнедеятельности, например прием пищи и поступление ее в желудок усиливают отделение желудочного сока, необходимого для гидролиза веществ. Появляющиеся в желудке и частично всасывающиеся в кровь продукты гидролиза в свою очередь стимулируют сокоотделение, что ускоряет и усиливает дальнейшее переваривание пищи. Однако положительная обратная связь нередко приводит систему в неустойчивое состояние, способствует формированию «порочных кругов».
Отрицательная обратная связь - выходной сигнал уменьшает входной. Отрицательные обратные связи способствуют сохранению устойчивого, стационарного состояния системы. Например, когда концентрация углекислого газа в организме человека увеличивается, лёгким приходит сигнал к увеличению их активности и выдыханию большего количество углекислого газа.
Регуляция по возмущению предусматривает использование самого возмущения для выработки, компенсирующего воздействия, в результате которого регулируемый показатель возвращается к исходному состоянию. Например, уменьшение парциального давления О2 в атмосферном воздухе при подъеме на высоту является возмущающим воздействием для системы дыхания, обеспечивающей оптимальное для метаболизма содержание кислорода в крови.
49. Постэмбриональный период онтогенеза, его периодизация. Основные процессы: рост, формирование дефинитивных структур, половое созревание, репродукция, старение.
С антропологической точки зрения в постэмбриональном периоде выделяют следующие стадии:
Новорожденность – от рождения до 10 дней. Происходит кормление молозивом. Начало ускорения роста в длину. Замечается уменьшения массы тела.
Грудной возраст – от 10 дней до 1 года. Питание «зрелым» молоком. Прорезывание первых молочных зубов (с 6 мес). Период максимальный интенсивности роста, начало формирования изгибов позвоночника, навыки сидения, стояния, первые шаги. Начало познавательного развития – осмотр и узнавание, «детская речь».
Раннее детство – 1-3 года. Завершается рост молочных зубов, падение интенсивности роста, узнавание картинок, фантазия, введение «я», взрослая речь
Первое детство – от 4 до 7лет. Часто бывает первый ростовой скачок. С 6 лет прорезываются постоянные зубы, в конце периода появляются первые черты полового диморфизма и начало половой самоидентификации, освоение понятий пространства и последовательности времени, основы этики и группового поведения.
Второе детство – 8-12 лет (м), 8-11 (ж). Прорезывание постоянных зубов, активный рост, начинается развитие вторичных половых признаков, формирование истинных волевых актов и формирование социальной адаптации.
Подростковый период – 13-16 (м), 12-15 (ж). Второй ростовой скачок. Завершается половое созревание, происходит крупнейший морфофункциональный сдвиг во всех органах и системах. Активное интеллектуально развитие, личная и эмоциональная нестабильность.
Юношеский возраст – 17-21 (м), 16-20 (ж). Завешается интенсивное развитие организма и его роста. Происходит стабилизация личности и форм мировоззрения.
Первый зрелый возраст – 22-35 (м), 21-35 (ж)
Второй зрелый возраст – 36-60 (м), 36-55 (ж). Отмечается относительная стабильность функционирование организма, завершается формирование типично мужских и типично женских принципов строения и психики. В конце периода завершается женский репродуктивный цикл, т. е. менопауза и климакс.
Пожилой возраст – 61-74 (м), 56-74 (ж).
Старческий возраст – 75-90. Продолжается оптимальная социальная активность. Начинаются старческие процессы в различных органах и системах, снижаются адаптационные способности организма. Завершается мужской репродуктивный период.
Долгожители – старше 90 лет. Относительная стабильность параметров организма на достигнутом уровне.
50. Старение как закономерный этап онтогенеза. Проявление старения на молекулярно-генетическом, клеточном, тканевом, органном и организменном уровнях.
Старение – это снижение жизнеспособности со временем, повышение вероятности смерти со временем. Классификация позднего онтогенеза:
Пожилой возраст 60-74
Старческий возраст 75-89
Долголетия 90+
Ведущими механизмами старости являются:
а) на молекулярном уровне: необратимые изменения ДНК, накапливающиеся в ходе онтогенеза, изменения в системе передачи генетической информации, изменения в синтезе РНК и белков разных классов, нарушения процессов преобразования, транспорта и использования энергии, снижение активности систем антиоксидантов, падение интенсивности синтеза гормонов и медиаторов;
б) на клеточном и субклеточном уровнях: деградация и гибель части клеток, снижение митотической активности клеток, уменьшение количества митохондрий, разрушение лизосом, изменение свойств (в том числе электрических) плазмолеммы, обезвоживание коллоидов цитоплазмы, накопление шлаков (например, пигмента липофусцина);
в) на органном и организменном уровнях: ослабление функции основных систем организма (нервной, эндокринной, сердечно-сосудистой, пищеварительной и др.), снижение нервного и гуморального контроля за их деятельностью, изменение чувствительности к действию гормонов.
Примеры изменений:
Седина
Снижение числа желез кожи и слизистых
Опорно-двигательный аппарат – развивается остеопороз (снижение плотности костной ткани), уменьшается слой хрящей в суставах и возникает артроз, сопровождающийся болью и нарушением функций.
Непереносимость молока
Мышцы – снижается тонус скелетной мускулатуры
Легкие – уменьшение числа альвеол на 30-50%, уменьшается жизненная емкость легких
ССС – снижение силы сокращения миокарда, что приводит к сердечной недостаточности, атеросклеротические повреждения сосудов.
Выделительная система – уменьшается количество нефронов, снижается почечный кровоток, снижается скорость клубочковой фильтрации
Нервная система – снижается количество нейронов, уменьшается толщина миелинового слоя, а значит замедляется проведение нервного импульса, повышается частотный порог восприятия звука.
Обмен веществ – снижается основной обмен, снижается чувствительность к инсулину и утилизации глюкозы.
Снижается работа иммунной системы.
51. Основные закономерности процесса старения. Гипотезы старения.
Закономерности старения:
Гетерохромность – возрастные изменения в различных органах начинаются в разном возрасте (тимус 15-18, яичники 48-52)
Гетеротопность – степень возрастных изменений неодинакова для разных органов и даже для разных участков одного органа (седина, тимус, пучковая зона коры надпочечников страдает меньше, чем клубочковая и сетчатая)
Гетерокинетичность – скорость возрастных изменений не одинакова в разных органах (костная ткань – небольшая скорость, нейроны ЦНС – высокая скорость).
На сегодняшний день имеется около 300 гипотез старения. Теории старения:
Гипотеза истощения (Рубнер) – организм при рождении получает запас энергии, когда энергия заканчивается, то организм гибнет.
Теория самоотравления (Мечников) – в кишечнике имеются гнилостные микробы, с возрастом количество микробов увеличивается. Эти микробы выделяют вредные вещества, которые отравляют человека и вызывают старение.
Теория Павлова – с возрастом снижается регуляторная активность нервной системы, и дисбаланс работы органов приводит к старению.
Молекулярно-генетические теории занимают доминирующее положение. Эти теории предполагают, что причиной старения и смерти является нарушение структурно-функциональной целостности наследственного аппарата.
Репарационная теория – с возрастом снижается активность ферментов репарации ДНК, однако спонтанные мутации происходят с обычной скоростью, поэтому в клетках накапливаются поврежденные ДНК, что нарушает синтез белка.
Теломеразная теория (Оловников) – при каждом клеточном делении хромосомы немного укорачиваются. У хромосом имеются особые концевые участки – теломеры, которые после каждого удвоения хромосом становятся немного короче, и в какой-то момент укорачиваются настолько, что клетка уже не может делиться. Тогда она постепенно теряет жизнеспособность – именно в этом, согласно теломерной теории, и состоит старение клеток.
Эффект Хейфлика – клетки здоровых тканей способны делиться определенное количество раз.
Теория феноптоза (Скулачев). Скулачевым выдвинуто предположение, о существовании некой генетической программы самоуничтожения, которая постепенно и разрушает организм. Самоубийство происходит не только на клеточном уровне- апоптоз, но и на субклеточном (разрушение органелл, например, митохондрий, при неправильном функционировании-митоптоз), органном- органоптоз, а самое главное, на организменном- феноптоз.
52. Смерть как биологическое явление (клиническая, биологическая). Социальная и биологическая составляющая здоровья и смертности в популяциях людей.
Смерть – это остановка жизненно важных функций. Однако организм сразу не погибает. Жизнь сохраняется 4-6 минут. Это определяется способностью головного мозга сохранять жизнеспособность без получения питательных веществ и кислорода. Этот период называется клинической смертью. Такая смерть может произойти при больших кровопотерях, электротравме. Признаки клинической смерти:
Отсутствие пульса
Отсутствие дыхания
Потеря сознания – отсутствие реакции на происходящие события, звуковые и болевые раздражители
Широкие зрачки и отсутствие их реакции на свет
Обнаружив клиническую смерть необходимо обеспечить адекватные реанимационные мероприятия – для этого достаточно наличие одного из первых двух признаков. Однако в дальнейшем происходит биологическая смерть. В этом случае происходят необратимые изменения головного мозга. Реанимация не нужна. Признаки биологической смерти:
Отсутствие реакции глаза на раздражение (надавливание)
Помутнение роговицы, образование треугольников высыхания
Появление симптома «кошачьего глаза»: при боковом сдавлении глазного яблока зрачок трансформируется в вертикальную веретенообразную щель, похожую на кошачий зрачок.
Снижение температуры тела
Трупные пятна
Трупное окоченение.
Социальная и биологическая составляющая здоровья и смертности в популяциях людей ????????????
53. Роль отечественных ученых в развитии общей и медицинской паразитологии (В.Н. Догель, В.Н. Беклемишев, Е.Н. Павловский, К.И. Скрябин).
Владимир Николаевич Беклемишев (1890-1962) – советский зоолог. Выполнены классические исследования биологии малярийного комара и других кровососущих членистоногих. Под его руководством была разработана система мер, в результате которых в СССР была почти полностью ликвидирована малярия. Он создал ряд оригинальных концепций в биоценологии и общей паразитологии. Работы В. Н. Беклемишева посвящены вопросам зоологии беспозвоночных, биоценологии, паразитологии и эпидемиологии. Крупный знаток низших ресничных червей. Беклемишев был ярчайшим представителем идеалистической морфологии, придающей главное значение плану строения организма.
Валентин Александрович Догель (1882-1955) – советский зоолог. Наиболее ценные его труды связаны с паразитологией протистологией. В. А. Догель уделял большое внимание работе в области сравнительной анатомии беспозвоночных животных, эмбриологии и разработке общебиологических вопросов. Создал концепции поли- и олигомеризации гомологичных органов в эволюции животных. Исследовал болезни рыб, в частности изучения природных очагов этих болезней. Исследования показали, что многие заражения человека паразитическими червями, передаются через рыбу, имеют очаговый характер. Издал монографию «Общая протистология».Во всех работах Догеля производится анализ всей паразитофауны исследованных животных, а не отдельных ее групп. Он рассматривает паразитофауну исследованных животных «как оригинальный биоценоз связанных цепью тесных взаимоотношений паразитических организмов, которых и следует, поэтому изучать в их совокупности, а не отдельно». Разработал основные вопросы взаимоотношения паразитических животных с внешней средой. Издал труд «Курс общей паразитологии», «Курс зоологии беспозвоночных», «Руководство по зоологии».
Евгений Никанорович Павловский (1884-1965) – советский зоолог, паразитолог. Под его руководством были организованы и проведены многочисленные комплексные экспедиции для изучения эндемичных, паразитарных и трансмиссивных заболеваний (клещевого возвратного тифа, клещевого энцефалита, москитной лихорадки, лейшманиозов и др. болезней). Е. Н. Павловским собраны обширные материалы по фауне, биологии и экологии паразитов и переносчиков болезней; изучены природные резервуары возбудителей болезней и пути их циркуляции в природе, а также в организме человека и сельскохозяйственных животных. Е. Н. Павловскому принадлежат исследования кишечно-протозойных и глистных инвазий, изучение фауны летающих кровососущих насекомых (гнус). Ученым разработаны способы защиты людей от этих насекомых (защитные сетки Павловского) и методы уничтожения кровососов в местах их размножения и обитания. Известны работы Е. Н. Павловского по изучению ядовитых животных и свойств их ядов. Им создано учение о природной очаговости трансмиссивных болезней человека, которое явилось основой развития экологического направления в паразитологии. Разработав учение о природной очаговости трансмиссивных болезней человека и сельскохозяйственных животных, заложил основы медицинской и ветеринарной биогеографии.
Константин Иванович Скрябин (1878-1972) – советский гельминтолог. К. И. Скрябин возглавлял первую в нашей высшей школе кафедру паразитологии в Донском ветеринарном институте. Он разработал метод полных гельминтологических вскрытий. В 1919 возглавляемая Скрябиным кафедра организовала первую гельминтологическую экспедицию. В конце 1920 года ученый организовал и возглавил гельминтологический отдел. Этот отдел был реорганизован во Всесоюзный институт гельминтологии. Скрябин организовал гельминтологический отдел Тропического института. Ученый руководил лабораторией АН СССР. Исследования Скрябина посвящены морфологии, биологии, филогении и систематике паразитических червей, вопросами эпидемиологии (эпизоотологии) и организации мероприятий по борьбе с гельминтозами человека и хозяйственно-полезных животных. Под руководством Скрябина проведено около 300 гельминтологических экспедиций в различные районы СССР. Он ввел понятие дополнительных резервуаров и транзитных хозяев и дал анализ процесса миграции различных гельминтов в теле хозяина; понятие гео- и биогельминтозов и др. Скрябин описал около 200 новых видов гельминтов. Он внес много нового в систематику гельминтов. Скрябиным была произведена ревизия и коренная перестройка всех классов паразитических червей. Он внес много нового и в разработку ряда общих биолого-гельминтологичеких проблем: изучение форм взаимоотношений между гельминтами, их хозяевами и внешней средой; определение жизненных циклов гельминтов, различных категорий хозяев и др. огромное практическое значение имеет разработанные Скрябиным методы дегельминтизации и девастации в борьбе с гельминтозами. Разработанные Скрябиным многочисленные инструкции и наставления по борьбе с гельминтозами были широко внедрены в медицинскую и ветеринарную практику.
54. Учение Е.Н. Павловского о природной очаговости болезней. Компоненты природного очага. Паразитарные природно-очаговые трансмиссивные и нетрансмиссивные заболевания, их критерии. Трансмиссивные болезни (облигатные и факультативные, антропонозы, зоонозы, антропозоонозы).
Большая группа паразитарных и инфекционных заболеваний характеризуется природной очаговостью. Для них характерны следующие признаки: 1) возбудители циркулируют в природе от одного животного к другому независимо от человека; 2) резервуаром возбудителя служат дикие животные; 3) болезни распространены не повсеместно, а но ограниченной территории с определенным ландшафтом, климатическими факторами и биогеоценозами. Компонентами природного очага являются: 1) возбудитель; 2) восприимчивые к возбудителю животные – резервуары; 3) соответствующий комплекс природно-климатических условий, в котором существует данный биогеоценоз. Особую группу природно-очаговых заболеваний составляют трансмиссивные болезни (то есть, заражение которыми через укус), такие, как лейшманиоз, трипаносомоз, клещевой энцефалит. Поэтому обязательным компонентом является также наличие переносчика. Категорию заболеваний с природной очаговостью выделил Павловский. Есть также нетрансмиссивные заболевания, они передаются при инъекциях, переливании крови, плазмы и т.п..
Трансмиссивные болезни:
Облигатные - трансмиссивные инфекционные болезни, возбудители которых передаются человеку или животному только через посредство специфических переносчиков из числа кровососущих членистоногих. К облигатно-трансмиссивным заболеваниям относят энцефалиты человека (клещевой и комариный), малярию, клещевые риккетсиозы (клещевой возвратный тиф, сыпной возвратный тиф, лихорадка паппатачи), лейшманиоз.
Факультативные – трансмиссивные заболевания, которые передаются как через переносчиков, так и через корма, воду в результате контакта с зараженным животным. К ним относят различные кишечные инфекции, сибирскую язву, туляремию.
Помимо такого деления, все инфекции, которыми заражается и болеет человек, принято разделять еще на две группы:
1. Антропонозы - заболевания, свойственные только человеку и передающиеся от человека человеку.
2. Зоонозы – болезни, свойственные животным и человеку и передающиеся от животного человеку, от человека человеку не передаются.
3. Антропозоонозы - группа инфекционных и инвазионных болезней, общих животным и человеку. К зооантропонозам относится сибирская язва, сап, бруцеллёз, туберкулёз, бешенство, ящур, лептоспироз, трипаносомоз, эхинококкоз, дифиллоботриоз и др.
55. Циклы развития паразитов. Чередование поколений в циклах развития паразитов (на примере представителей разных типов и классов)??????. Понятие об основных, резервуарных и промежуточных хозяевах.
Онтогенез паразитов обычно бывает сложнее, чем развитие свободноживущих видов. Поэтому большинство паразитов нередко развиваются со сложным метаморфозом, включающим много личиночных стадий, обитающих в разных средах и выполняющих разные функции: расселение, активного роста, пассивного ожидания попадания в другую среду обитания и иногда даже размножения. Совокупность всех стадий онтогенеза паразита и путей передачи его от одного хозяина к другому называют его жизненным циклом. Личинки могут вести как свободный, так и паразитический образ жизни. Хозяин, в котором обитают личинки паразита, носит название промежуточного. Значение промежуточных хозяев в циклах развития паразитов очень велико: они являются источниками заражения окончательных хозяев, часто выполняют расселительные функции. Иногда в цикле развития паразита последовательно сменяются два, три промежуточных хозяина. Хозяина, в котором развивается и размножается половым путем половозрелая стадия паразита, называют окончательным или дефинитивным. Заражение его осуществляется либо при поедании промежуточного хозяина, либо при контакте с последним в одной среде обитания. Выделяют также понятие «резервуар паразита», или «резервуарный хозяин». Это такой хозяин, в организме которого возбудитель заболевания может жить долго, накапливаясь, размножаясь и расселяясь по окружающей территории.
56. Подцарство Простейшие. Систематическое положение и классификация подцарства. Характерные черты организации. Представители, имеющие медицинское значение.
Систематика:
Империя – клеточные
Надцарство – эукариот
Подцарство – одноклеточные или простейшие (Protozoa)
Типы 1) Саркомастигофоры
Подтип Саркодовые (Sarcodina)
Класс – корненожки (Rhizopoda)
Отряд – амебы (Amoebina)
Род Entamoeba hystolitica, gingivalis, coli.
Подтип Мастигофоры – жгутиковые
Класс – зоомастигина
Отряд – одножгутиковые (Protomandina)
Род – трипаносома (3)
Род – лейшмании (5)
Отряд - многжгутиковые
Род – Lamblia intestinalis
Род – Trichomonas hominis, vaginalis.
2) Инфузории
Род – Balantidium coli
3) Апикомплекс
Класс – Sporozoa
Отряд - кровяные
Род – plasmodium (4)
Отряд – кокцидии
Род – токсоплазма
Общая характеристика. Тело состоит из цитоплазмы, ограниченной наружной мембраной, ядра и органеллы обеспечивают питание, движение, выделение. Питание может осуществляться разными способами: 1. Фагоцитоз – частица пищи обтекается и оказывается внутри вакуоля; 2. Пиноцитоз – всасывание поверхностью тела. Питаются простейшие органическими частицами, микроорганизмами. Большинство имеет органы передвижения: жгутики, реснички, псевдоподии. Размножение осуществляется разными способами: а) митоз; б) копуляция – слияние клеток; в) конъюгация – обмен частью наследственного материала. В жизненном цикле существует трофозоита (активно питающаяся и перемещающаяся форма) и циста (неподвижная форма, покрытая плотной оболочкой и с медленным обменом веществ).
57. Дизентерийная амеба. Систематическое положение, морфология, цикл развития, пути заражения, обоснование методов лабораторной диагностики и мер профилактики. (Систематика см 56)
Дизентерийная амеба (Entamoeba histolitica) – возбудитель амебиаза. Имеется несколько стадий. Мелкая вегетативная форма (8-20 мкм) обитает в просвете кишки. Крупная форма (45 мкм) в просвете кишки в гнойном содержании язв стенки. Цитоплазма четко разделена на прозрачную, стекловидную и зернистую. В ней располагается ядро и эритроцит, которым она питается.
Жизненный цикл: человек заражается амебиазом, проглатывая цисты паразита. В просвете толстой кишки из цисты образуется 4 мелкие вегетативные формы. Далее мелкие могут превращаться в крупные, вызывая образование язв. В тяжелых случаях они превращаются в тканевые формы, выходя в кровь.
Методы диагностики – обнаружение трофозоитов в фекалиях
Профилактика – соблюдение гигиены питания.
58. Лямблия, трихомонады. Систематическое положение (см 56), морфология, цикл развития, пути заражения, обоснование методов лабораторной диагностики и мер профилактики.
Лямблии (Lamblia intestinalis). Морфология. Лямблии существуют в виде вегетативной формы и способны образовывать цисты. Вегетативная форма активная, подвижная, грушевидной формы. Передний конец тела закруглен, задний заострен. Длина 9-18 мкм. В передней части тела находится присасывательный диск в виде углубления. Имеет 2 ядра, 4 пары жгутиков. Джижение за счет бокового переворачивания вокруг продольной оси. Пищу всасывают всей поверхностью тела.
Жизненный цикл. Лямблии обитают в верхнем отделе тонкого кишечника. С помощью присасывательного диска прикрепляются к ворсинкам. И попадая в нижние отделы кишечника, превращаются в цисты, которые выделяются с испражнениями. Цисты хорошо сохраняются в окружающей среде. Заражение через грязные руки, игрушки, пищу и воду. Цисты, попадая в кишечник, превращаются там в вегетативные формы. Одна циста образует две вегетативные формы.
Методы диагностики – обнаружение цист в фекалиях.
Профилактика – личная гигиена.
Трихомонады. В организме человека обитает три вида трихомонад: Trichomonas hominis – кишечная – в толстом кишечнике;Trichomonas tenax – ротовая в полости рта; Trichomonas vaginalis – мочеполовая – в мочеполовых путях.
Кишечная трихомонада. Морфология. Имеет грушевидное тело длиной 8-20 мкм. От переднего конца тела отходят обычно 5 жгутиков. С одной стороны тела по всей его длине расположена ундулирующпая мембрана. Есть осевая нить – аксостиль.
Жизненный цикл. Размножается путем деления цист не образует.
Методы диагностики – обнаружение паразитов в фекалиях.
Профилактика – личная гигена.
Мочеполовая (влагалищная) трихомонада. Морфология. Имеет грушевидное тело длиной 14-30 мкм. На переднем конце тела расположены 4 жгутика и ундулирующая мембрана. Сквозь все тело проходит осевая нить, выступающая на заднем конце в идее шипика.
Жизненный цикл – цист не образует, передается половым путем.
Методы диагностики – обнаружение трихомонад в выделениях мочеполовых путей и моче.
59. Лейшмании. Систематическое положение (см 56), морфология, цикл развития, пути заражения, локализация в организме человека, патогенное действие, обоснование методов лабораторной диагностики и мер профилактики.
Для человека патогенны несколько видов лейшманий: Leishmania tropica , вызывает антропонозный (городской) кожный лейшманиоз; Leishmania major — возбудитель зоонозного (пустынного) кожного лейшманиозa; Leishmania braziliensis — кожно-слизистый (американский) лейшманиоз; Leishmania donovani — вызывает висцеральный, или внутренний, лейшманиоз; Leishmania infantum — возбудитель висцерального лейшманиоза.
Морфология. Лейшмании проходят две стадии развития: безжгу-тиковую и жгутиковую. Безжгутиковая форма овальная, длиной 2—6 мкм. Ядро округлое, занимает до '/з клетки. Жгутиковая форма подвижная, жгутик длиной 15— 20 мкм. Тело удлиненное веретенообразное, длиной до 10—20 мкм. Деление продольное. Развиваются в теле переносчика - москита.
Жизненный цикл. Лейшманиозы входят в группу трансмиссивных инфекций, переносчиками служат мелкие кровососущие насекомые — москиты. Москиты заражаются при кровососании на больных людях или животных. В первые же сутки заглоченные безжгутиковые паразиты превращаются в подвижные жгутиковые формы, начинают размножаться и спустя 6— 8 дней скапливаются в глотке москита.
При укусе человека или животного зараженным москитом подвижные лейшмании из его глотки проникают в ранку и затем внедряются в клетки кожи или внутренних органов в зависимости от вида лейшмании. Здесь происходит их превращение в безжгутиковые формы.
Патогенное действие. Висцеральным (средиземноморским) лейшманиозом чаще заболевают дети. После инкубационного периода длительностью от нескольких недель до нескольких месяцев у заболевшего повышается температура тела, появляются вялость, адинамия, бледность, исчезает аппетит. Увеличиваются селезенка и печень, вследствие чего заметно выступает живот . Развиваются анемия и истощение больного. Заболевание тянется несколько месяцев и при отсутствии специфического лечения обычно заканчивается смертью.
При кожном лейшманиозе после инкубационного периода (1—2 мес) в местах укусов москитов появляются небольшие бугорки буровато-красноватого цвета, средней плотности, обычно малоболезненные. Бугорки постепенно увеличиваются и затем изъязвляются через 3—б нед при антропонозной форме и через 1—3 нед при зоонозной. Возникают язвы с отеком окружающей ткани, воспалением и увеличением лимфатических узлов. Процесс тянется несколько месяцев, при антропонозной форме — больше года, заканчиваясь выздоровлением. На месте язв остаются рубцы. После перенесенного заболевания формируется стойкий иммунитет.
Метод диагностики. Окончательный диагноз висцерального лейшманиоз а ставят на основании обнаружения лейшмании при микроскопии мазков костного мозга. При кожном лейшманиозе материал получают поскабяиванием скальпелем нераспавшихся бугорков или инфильтрата по краю язв до появления серозно-кровянистой жидкости.
Профилактика. С целью предупреждения заболеваемости всеми видами лей-шманиозов проводят уничтожение москитов, ликвидацию мест выплода, обработку мест пребывания ядохимикатами, а также принимают меры по предохранению людей от укусов москитов.
60. Трипаносомы. Систематическое положение (см 56), морфология, цикл развития, пути заражения, обоснование методов лабораторной диагностики и мер профилактики.
Из рода трипаносом для человека патогенны три вида: Trypanosoma gambiense и Trypanosoma rhodesiense вызывают африканский трипаносомоз (сонную болезнь) и Trypanosoma cruzi — возбудитель американского трипаносомоза (болезнь Шагаса).
Морфология. Тело трипаносом продолговатое, узкое, имеет жгутики и ундулирующую мембрану. Длина тела трипаносом 17—28 мкм, в средней части находится овальное ядро.
Жизненный цикл. Первая часть жизненного цикла проходит в пищеварительном канале мухи цеце, вторая — в организме хозяина. У позвоночных животных трипаносома находится в крови, откуда при укусе попадает в желудок мухи. Переносчик африканского трипаносомоза кусает следующего хозяина. Она поселяется в крови, лимфе, спинномозговой жидкости.
Методы диагностики - мазок крови, спинномозговая жидкость.
Профилактика. Личная — прием профилактических лекарственных препаратов. Общественная — уничтожение переносчиков и мест их выплода.
61. Балантидий. Систематическое положение (см 56), морфология, цикл развития, пути заражения, обоснование методов лабораторной диагностики и мер профилактики.
Морфология. Балантидий (Balantidium coli) — самый крупный представитель паразитических простейших человека. Вегетативная форма вытянутая, чаще яйцеобразная. Длина 30—150 мкм, ширина 30—100 мкм. С помощью многочисленных ресничек балантидий активно двигаются, нередко вращаясь при этом вокруг своей оси. Питаются различными пищевыми частицами, включая бактерии, грибы, форменные элементы крови, для заглатывания которых служит цитостом (клеточный рот). Цитоплазма содержит пищеварительные и две пульсирующие выделительные вакуоли.
Жизненный цикл. Балантидии обитают в кишечнике свиней и с испражнениями попадают в окружающую среду. И с загрязненной водой или пищей цисты попадают в толстый кишечник человека.
Методы диагностики. Обнаружение балантидиев в фекалиях
Профилактика. Соблюдение правил личной гигиены, особенно при уходе за свиньями. Охрана от загрязнения воды и пиши.
62. Малярийный плазмодий. Систематическое положение (см 56), морфология, цикл развития, пути заражения, обоснование методов лабораторной диагностики и мер профилактики.
Плазмодии, вызывающие у человека малярию, относятся к четырем видам: Plasmodium vivax — возбудитель трехдневной малярии; Plasmodium malariae — возбудитель четырехдневной малярии; Plasmodium falciparum — возбудитель тропической малярии и Plasmodium ovale — возбудитель малярии, типа трехдневной (овале).
Жизненный цикл. Малярийные плазмодии проходят две стадии развития — бесполую (шизогонию) в организме человека и половую (спорогонию) в организме переносчика — самок малярийных комаров рода Anopheles.
При кровососании зараженный малярийный комар вместе со слюной вводит в ранку спорозоиты . С кровью они попадают в клетки печени, где развиваются и делятся. Образовавшиеся в результате деления в клетках печени молодые паразиты (мерозоиты) поступают в кровь и проникают в эритроциты — наступает эритроцитарная шизогония.
Мерозоиты, проникшие в эритроциты, превращаются в трофозоиты (растущие формы), а последние — в шизонты (делящиеся формы). Указанный цикл развития в эритроцитах составляет 72 ч для P. malariae или 48 ч для остальных видов.
В некоторых эритроцитах развиваются мужские и женские половые формы — гамонты. Они завершают свое развитие, только попав в организм комара с кровью. В результате в слюнных железах малярийного комара скапливаются спорозоиты, и такой комар становится способным вновь заражать людей.
Методы диагностики. Обнаружение паразитов вмазке крови.
Профилактика. Ликвидация малярийных комаров.
63. Токсоплазма. Систематическое положение, морфология, цикл развития, пути заражения, обоснование методов лабораторной диагностики и мер профилактики.
Морфология. Токсоплазма (Toxoplasma gondii) по форме тела напоминает полумесяц. Один конец заострен, другой — закруглен. Длина 4—7 мкм.
Жизненный цикл. Половой цикл размножения происходит в клетках слизистой оболочки кишечника кошек (основной хозяин). С испражнениями кошек выделяются ооцисты. Ооцисты созревают и хорошо сохраняются в почве, при их заглатывании заражаются животные, в том числе грызуны. Человек заражается токсоплазмами: 1) через рот (алиментарный, или пищевой путь) при употреблении сырого или полусырого мяса, фарша, а чаще — через загрязненные ооцистами (от кошек) овощи, ягоды, предметы, руки; 2) иногда через кожу рук и слизистые оболочки, например при разделке мясных туш, лабораторных работах с заразным материалом; 3) внутриутробно (трансплацентарный путь). Токсоплазма паразитирует в клетках печени, легких, лимфоидной ткани.
Методы диагностики. Исследование крови, спинномозговой жидкости, пунктатов лимфатических узлов, плаценты. Возможна внутрикожная аллергическая проба с токсоплазмином.
Профилактика. Для профилактики всех форм токсоплазмоза важно соблюдать правила содержания кошек. Особенно тщательно меры предосторожности необходимо выполнять беременным женщинам, а также медицинскому персоналу при работе с инфицированным материалом.
64.Токсоплазма — возбудитель токсоплазмоза. Имеет форму полумесяца, один конец которого заострен более другого. В центре располагается крупное ядро. Дина паразита 4-7 мкм. Жизненный цикл токсоплазмы типичен для споровиков: в нем чередуются стадии шизогонии, гаметогонии и спорогонии. Основные хозяева паразита — домашние кошки и дикие виды семейства Кошачьи. Они заражаются, поедая больных грызунов, птиц или инвазированное мясо крупных животных. Паразиты у них сосредотачиваются в клетках кишечника, размножаются шизогонией, а затем образуют гаметы. После копуляции гамет формируются ооцисты, которые выделяются во внешнюю среду. В них происходит спорогония, т.е. деление зиготы под оболочкой.
Своеобразной особенностью цикла развития токсоплазм является то, что промежуточные хозяева могут заражаться ими не только от основного хозяина, но и при поедании друг друга. Возможно и внутриутробное заражение плода от больной беременной самки, когда паразиты проникают через плаценту.
В соответствии с этим и человек как промежуточный хозяин может заразиться токсоплазмозом разными путями: 1) при поедании мяса инвазированных животных; 2) с молоком и молочными продуктами; 3) через кожу и слизистые оболочки; 4) внутриутробно через плаценту; 5) при медицинских манипуляциях переливания крови и лейкоцитарной массы, при пересадках органов, сопровождающихся приемом иммунодепрессивных препаратов.
Наиболее опасным является трансплацентарное заражение. При этом возможно рождение детей с множественными врожденными пороками развития, в первую очередь головного мозга. Для исследования используют плаценту, печень, кровь, лимфатические узлы, головной мозг.
Профилактика — термическая обработка животных продуктов питания, санитарный контроль на бойнях и мясокомбинатах, предотвращение тесных контактов детей и беременных женщин с домашними животными.
65. Сосальщики - это паразитические черви с плоским листовидным или удлиненным нерасчлененным телом. Оно покрыто плотной защитной оболочкой; ресничного эпителия нет. К телу хозяина черви прикрепляются с помощью присосок. Тип плоские черви, класс сосальщики.
Сосальщики (Trematodes) — паразитические организмы. Известно около 3000 видов сосальщиков. Для этих паразитов характерны сложные циклы развития, в которых происходит чередование поколений, а также способов размножения и хозяев.
Половозрелая особь имеет листовидную форму. Рот расположен на терминальном конце тела и снабжен мощной мускулистой присоской. Кроме нее, имеется еще одна присоска на брюшной стороне. Дополнительными органами прикрепления у некоторых видов — мелкие шипики, покрывающее все тело.
Пищеварительная система мелких видов сосальщиков представляет собой мешок или два слепо заканчивающихся канала. У крупных видов она сильно разветвляется. Помимо функции собственно пищеварения, она выполняет еще и транспортную роль — перераспределяет продукты питания по всему телу. У плоских червей, в том числе у сосальщиков, отсутствует внутренняя полость тела, а значит, нет кровеносной системы. Листовидная форма тела дает возможность кишке снабжать все тело питательными веществами. Та же форма делает возможным газообмен через всю поверхность тела, поскольку органов и тканей, лежащих глубоко под кутикулой, просто нет.
Сосальщики — гермафродиты. Мужская половая система: пара семенников, два семяпровода, семяизвергательный канал, копулятивный орган (циррус). У печеночного сосальщика семенники ветвящиеся, у кошачьего и ланцетовидного — компактные. Женская половая система: яичник, яйцеводы, желточники, семяприемник, матка, половая клоака. Желточники обеспечивают яйцо питательными веществами, скорлуповые железы — оболочками. Осеменение внутреннее, перекрестное. Яйца созревают в матке.
Половозрелая особь (марита) всегда обитает в организме позвоночного животного. Она выделяет яйца. Для дальнейшего развития яйцо должно попасть в воду, где из него выходит личинка — мирацидий. Личинка имеет светочувствительные глазки и реснички, способна самостоятельно отыскивать промежуточного хозяина, используя различные виды таксиса. Мирацидий должен попасть в организм брюхоногого моллюска, строго специфичного для данного вида паразита. В его организме личинка превращается в материнскую спороцисту, которая претерпевает
наиболее глубокую дегенерацию. Она имеет только женские половые органы, поэтому и размножается только партеногенетически.
При ее размножении формируются многоклеточные редии, которые также размножаются партеногенезом. Последнее поколение редий может генерировать церкарии. Они покидает организм моллюска и для дальнейшего развития должны попасть в тело окончательного или второго промежуточного хозяина. В первом случае церкарии либо активно внедряются в организм окончательного хозяина, либо инцистируются на траве и заглатываются с нею.
Во втором случае церкарии ищут тех животных, которые используются основным хозяином в пищу, и формируют в их теле покоящиеся стадии — инцистированные метацеркарии. Основная масса церкариев погибает, не попав в организм основного хозяина, так как они неспособны к активному поиску, либо попадают в организм тех видов, развитие в которых невозможно. Способность паразита размножаться на личиночных стадиях значительно увеличивает его популяцию.
После проникновения в организм окончательного хозяина инвазионные стадии сосальщиков мигрируют в нем и находят нужный для дальнейшего развития орган. Там они достигают половой зрелости и обитают.
Миграция по организму сопровождается явлениями тяжелой интоксикации и аллергическими проявлениями.
Заболевания, вызываемые сосальщиками, носят общее название трематодозов.
66.Печеночный сосальщик - листовидное тело от 3 до 5 см с двумя присосками - ротовой и брюшной. На дне ротовой присоски находится рот, ведущий в глотку, от которой отходят 2 ветви средней кишки с многочисленными боковыми выростами. В жизненном цикле наблюдается смена хозяев. Взрослая гермафродитная особь живет в печени окончательного (основного) хозяина (овцы, крупного рогатого скота, свиньи, лошади, изредка человека). Питается в основном желчью, вызывая истощение или даже гибель хозяина. Оплодотворенные яйца паразита вместе с желчью поступает в кишечник хозяина, выводятся во внешнюю среду. В пресном водоеме, где через 15-25 дней из яйца выходит личинка, с помощью которых она плавает. В теле промежуточного хозяина - моллюска малого прудовика личинка внедряется с помощью буравящего стилета, проникает в печень. Образуется спороциста, в ней образуется личинка. Личинка в теле промежуточного хозяина дважды размножается партеногенетически. Покинув тело моллюска, личинка свободно плавает, прикрепляется к растениям, теряет хвост, одевается плотной оболочкой и превращается в цисту, внутри которой сохраняет свою жизненную способность. С зеленым кормом цисты попадают в организм домашних животных, первоначально в желудок, оболочка цисты растворяется и паразит проникает в кишечник, из него в желчные протоки печени, вырастает во взрослого сосальщика. Человек заражается ими при употреблении сырой воды из водоема, овощей и фруктов, вымытых в этой воде.
Ряд приспособлений: форма тела, размеры, покров (покрыт плотной оболочкой, выполняющей защитную роль от действия пищеварительных соков хозяина); присоски (ими червь прикрепляется к печени хозяина), продукты жизнедеятельности (ядовитые жидкие вещества) выделяются в организм хозяина, вызывая страшные страдания животных и человека от заболевания; наличие в жизненном цикле паразита кроме размножающегося половым путем гермафродитного поколения еще и партеногенетического размножения (без оплодотворения) у личинок можно рассматривать как приспособительный признак, обеспечивающий резкое повышение интенсивного размера.
Профилактика - уничтожение в местных водоемах малого прудовика (промежуточного хозяина) химическими средствами, лечебными препаратами - с взрослой формой этого паразита.
Диагностика — обнаружение яиц в фекалиях, моче или мокроте, аллергические пробы.
67. Кошачий, или сибирский, сосальщик (Opisthorchis felineus) – возбудитель описторхоза.
Кошачий сосальщик имеет бледно-желтый цвет, длина его – 4—13 мм. В средней части тела находится разветвленная матка, за ней – округлый яичник. Характерная особенность – наличие в задней части тела двух розетковидных семенников, которые хорошо окрашиваются.
Окончательные хозяева паразита – дикие и домашние млекопитающие и человек. Первый промежуточный хозяин – моллюск Bithinia leachi. Второй промежуточный хозяин – карповые рыбы, в мышцах которых локализуются метацеркарии.
Сначала яйцо с мирацидием попадает в воду. Далее оно заглатывается моллюском, в задней кишке которого мирацидий выходит из яйца, проникает в печень и превращается в спороцисту. В ней путем партеногенеза развиваются многочисленные поколения редий, из них – церкарии. Церкарии попадают в воду и, активно плавая в ней, внедряются в тело рыбы или заглатываются ею. Эта стадия развития называется метацер-карием. При поедании окончательным хозяином сырой или вяленой рыбы метацеркарии попадают в его желудочно-кишечный тракт. Под влиянием ферментов оболочки растворяются. Паразит проникает в печень и желчный пузырь и достигает половой зрелости.
Диагностика. Обнаружение яиц кошачьего сосальщика в фекалиях и дуоденальном содержимом, полученном от больного.
Профилактика. Соблюдение правил личной гигиены. Санитарно-просветительская работа.
Миллионы людей поражены описторхозом и клонорхозом в странах Юго-Восточной Азии. В Таиланде описторхисами инвазированы 80% населения северных районов. В России и странах СНГ основные очаги описторхоза находятся в бассейнах рек Оби, Иртыша, Волги, Камы, Днепра, Северной Двины, Немана, Днестра, Южного Буга и рек Крайнего севера России и Казахстана. Высокому распространению описторхоза способствуют некоторые географические и социально-бытовые условия Пермской области.
68.Китайский сосальщик.
Гельминтоз из группы трематодозов, поражающий печень и поджелудочную железу. Возбудителем заболевания является сосальщик «Clonorchis sinensis» - клонорхоз китайский, размеры его варьируют от 10 до 20 мм. Клонорхоз широко распространен в Китае, Японии, на Корейском полуострове. В России встречается в бассейне реки Амур. Зрелые формы паразитируют у человека и рыбоядных млекопитающих.
|
|
Источники
Возбудителем клонорхоза является трематода семейства Opisthorchidae — Clonorchis sinensis (синоним — двуустка китайская). Впервые описан Мс Соnnеll в 1874 г., подробно изучен Kobajashi в 1910 г. Тело плоское, длиной 10—20 мм, шириной 2—4 мм. На переднем конце расположена ротовая присоска, на границе первой и второй четверти тела — брюшная присоска. Яйца желтовато-коричневого цвета с крышечкой на одном конце и утолщением скорлупы на противоположном; их размеры — 0,026-0,035 х 0,012-0,0195 мм, отличаются отсутствием шифтковидного выступа от яиц возбудителей описторхоза. Гельминт в стадии половой зрелости паразитирует у человека и плотоядных млекопитающих, которые являются дефинитивными хозяевами. Промежуточными хозяевами служат пресноводные моллюски, дополнительными — карповые рыбы и пресноводные раки. Длительность жизни возбудителя клонорхоза в организме человека до 40 лет.
Промежуточных хозяев у клонорха два: первый моллюски семейства «Bithyniiadae», и второй – рыбы семейства карповых, которые и являются источником заболевания для человека.
Пути заражения Заражения человека и животных происходит при употреблении в пищу сырой, мороженой, малосоленой и недостаточно термически обработанной инвазированной рыбы и моллюсков.
Так же источниками заражения являются инфицированные люди (в основном), кошки, собаки. Яйца гельминта, выделяемые с фекалиями, при попадании в воду заглатываются моллюсками, в теле которых примерно через 2 недели формируются личинки — церкарии. Церкарии активно проникают в подкожную клетчатку и мышцы рыб и раков, где превращаются в метацеркариев. Человек заражается при употреблении в пищу недостаточно термически обработанных инфицированных рыб и раков. Клонорхоз широко распространен в Китае, Корее, Японии, в бассейне Амура и Приморье.
Формы существования
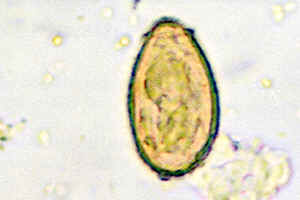 Метацеркарий
клонорха, попадая в кишечно-желудочный
тракт человека, освобождается от своей
оболочки и проникает в желчный пузырь
и поджелудочнуюжелезу, где и обитает.
Яйца паразита с желчью и поджелудочным
соком поступают в кишечник и выделяются
с фекалиями. Попавшие в воду яйца паразита
заглатываются рыбой и моллюсками.
Человек съедает зараженную рыбу, и цикл
распространения клонорха продолжается.
Срок жизни паразита составляет 25 лет и
более.
Метацеркарий
клонорха, попадая в кишечно-желудочный
тракт человека, освобождается от своей
оболочки и проникает в желчный пузырь
и поджелудочнуюжелезу, где и обитает.
Яйца паразита с желчью и поджелудочным
соком поступают в кишечник и выделяются
с фекалиями. Попавшие в воду яйца паразита
заглатываются рыбой и моллюсками.
Человек съедает зараженную рыбу, и цикл
распространения клонорха продолжается.
Срок жизни паразита составляет 25 лет и
более.
Симптомы клонорхоза
В острой стадии, через 2-4 недели после инвазии, возникает лихорадка, появляется сыпь, повышается уровень эозинофилов в крови, что определяется токсическим и аллергизирующим действием ферментов и продуктов жизнедеятельности личинок. Появляется кашель, боль в груди, желтуха. Длительность острой стадии 1-6, чаще 2-4 недели.
Хроническая стадия характеризуется поражением желчевыводящих путей, при этом снижается тонус желчного пузыря, возникает спазм и отек его протоков, нарушается поступление желчи в кишечник (холестаз) и работа поджелудочной железы. Возникают боли в области правого подреберья в эпигастрии и по ходу кишечника, тошнота, вздутие живота.
Последствия паразитирования в организме Первым объектом заселения чаще оказывается печень.Клонорх, как и все сосальщики, встраивается в ткани печени, фильтрующие кровь, и питается ею. При этом развивается общая интоксикация организма, растет уровень билирубина в крови, снижается белок крови, появляются дистрофические изменения миокарда, миокардит, инфильтраты в легких, развивается пневмония. Методы диагностики
Диагноз острого клонорхоза устанавливают на основании клинической картины, данных эпидемиологического анамнеза (например, пребывание в регионах распространения клонорхоза), подтверждают с помощью паразитологического исследования на яйца гельминта в фекалиях, дуоденальном содержимом (возможно обнаружить через 1-1,5 месяца после инвазии).
69. ЛЕГОЧНЫЙ СОСАЛЬЩИК Paragonimus westermani - биогельминт, возбудитель парагонимоза. Распространен в Юго-Восточной и Южной Азии, Центральной Африке и Южной Америке. Морфологические особенности. Форма тела яйцевидная, слегка сплющенная в дорзо-вентральном направлении; длина - 7,5-12 мм. Марита имеет красно-коричневую окраску. Ротовая присоска расположена терминальнобрюшная - примерно на середине тела. Каналы средней кишки неразветвленные, по ходу образуют изгибы. По бокам от брюшной присоски лежат с одной стороны дольчатый яичник, а с другой - матка. Желточники расположены в боковых частях тела. Кзади от матки и яичника лежат два лопастных семенника (рис. 2). Цикл развития типичен для трематод. Основными хозяевами легочного I сосальщика являются человек, собака, кошка, свинья и другие млекопитающие. Первый промежуточный хозяин - пресноводные моллюски рода Melania, второй - пресноводные раки и крабы. Заражение основного хозяина происходит при употреблении в пищу раков и крабов, не прошедших достаточную термическую обработку, в которых находятся живые метацеркарии. В желудочно-кишечном тракте паразиты освобождаются от оболочек, проникают через стенку кишечника в брюшную полость, а оттуда через диафрагму - в плевру и легкие. Локализация мариты - мелкие бронхи. Яйца выделяются во внешнюю среду с мокротой или фекалиями.
|
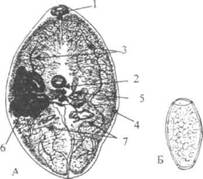
Рис. 2. Легочный сосалыцик (Paragonimus westermani). А - половозрелая форма. Б -яйцо. 1 - ротовая присоска; 2 - брюшная присоска; 3 -кишечник; 4 - желточники; 5 - яичник; 6 - матка; 7 - семенники. Патогенное действие заключается в механическом повреждении стенки кишечника, диафрагмы, плевры и ткани легких, в которых наблюдаются кровоизлияния и воспалительные процессы. Токсико-аллергическое действие проявляется лихорадкой и эозинофилией. В легких паразиты располагаются попарно; вокруг них образуются полости, заполненные продуктами обмена паразита и распада окружающих тканей. Яйца паразита с током крови могут заноситься в различные органы. Особую опасность представляет попадание яиц в головной мозг. Клиника. Инкубационный период - 2-3 недели, он может укорачиваться до нескольких дней при массивной инвазии. Ранние проявления заболевания характеризуются симптомами энтерита, гепатита, асептического перитонита, что связано с миграцией личинок. Эти явления длятся недолго. Позже развивается плевролегочной процесс, обусловленный паразитированием молодых гельминтов, что проявляется симптомами бронхита, очаговой пневмонии и, нередко, эксудативного плеврита. В этой стадии яйца парагонимусов в мокроте не обнаруживаются. Хроническая стадия, наступающая спустя 2-3 месяца после заражения, характеризуется интоксикацией с температурой до 39°С, кашлем с гнойной мокротой (до 500 мл в сутки) и нередко с примесью крови, болями в груди, одышкой, легочными кровотечениями. В крови - умеренный лейкоцитоз, эозинофилия. В мокроте находят яйца паразита. В дальнейшем происходит фиброз и кальцификация очагов, которые отчетливо видны на рентгенограммах. В нелеченных случаях развивается пневмосклероз и симптомокомплекс легочного сердца. При попадании яиц или взрослых гельминтов в головной мозг развиваются симптомы абсцесса мозга и менингоэнцефалита. Лабораторная диагностика основана на нахождении яиц в мокроте или фекалиях. Яйца парагонима относительно крупные (до 100 мкм), овальные, желтоватой окраски, с крышечкой и толстой оболочкой. Лечение. Высокоэффективным препаратом при парагонимозе является Bitionol (битионол). Его применяют внутрь по 30-40 мг/кг в сутки за 2 приема после еды. Препарат принимают через день на протяжении 2-х недель. Курсовая доза для взрослого - 300-400 мг/кг. Кроме битионола можно назначать Praziquantel (празиквантель) внутрь по 25 мг/кг 3 раза в день в течение 2-х суток. При своевременном лечении и низкой интенсивности инвазии прогноз благоприятный, при массивной инвазии часто развивается пневмосклероз, при поражении головного мозга прогноз сомнительный. Профилактика: личная - не употреблять в пищу сырых или плохо термически обработанных раков и крабов; общественная - санитарно-просветительная работа, охрана водоемов от загрязнения фекалиями человека и животных, выявление и лечение больных. 70. Шистосомо́зы (schistosomoses; синоним: шистосоматозы, шистосомиазы, бильгарциозы)
группа тропических гельминтозов, вызываемых плоскими червями-сосальщиками; характеризуются хроническим течением, поражением мочеполовой системы, кишечника, печени и других органов.
Различают мочеполовой Ш. (возбудитель — Schistosoma haematobium), кишечный шистосомоз Мансона (возбудитель — S. mansoni), японский Ш. (возбудитель — S. Japonicum) и кишечный интеркалатный (возбудитель — S. intercalatum). Наиболее широкое распространение имеет мочеполовой шистосомоз. Ареал Ш. охватывает страны тропического и субтропического пояса (Африка, Юго-Восточная Азия, Ближний Восток, Латинская Америка). Шистосомы раздельнополы. Морфология всех шистосом сходна. У самца широкое удлиненное тело, на брюшной стороне имеется гинекоформный канал, где в период копуляции помещается самка. Яйца овальные, имеют шип, расположенный, в зависимости от вида, сбоку или на одном из полюсов. Шистосомозы перкутантные биогельминтозы.
Жизненный цикл возбудителя характеризуется сменой двух хозяев (рис.). Источником возбудителя инвазии при всех Ш. является человек, а при японском, кроме того, домашние и дикие животные (коровы, козы, лошади, свиньи, собаки, кошки, грызуны), при меконговом — собаки. В мелких кровеносных сосудах окончательного хозяина — человека, многих млекопитающих самка откладывает до 3000 яиц в сутки. Яйца проходят в кишечник или мочевой пузырь и испражнениями или мочой выводятся в окружающую среду. В пресной воде из яйца вылупляются мирацидии. Дальнейшее их развитие происходит в организме промежуточного хозяина — моллюска, специфического для каждого вида шистосом. Через 4 нед. после внедрения в теле моллюска созревают хвостатые личинки — церкарии. Они выходят в воду, где при встрече с окончательным хозяином внедряются через кожу или слизистые оболочки. В подкожной клетчатке церкарии превращаются в шистосомулы, которые попадают в кровеносные сосуды и через 20—30 дней достигают половой зрелости.
Патогенез и патологическая анатомия. Мигрируя по кровеносным и лимфатическим сосудам, шистосомулы проникают в различные органы, вызывая их поражения. В сосудах портальной системы печени шистосомулы созревают до взрослых особей. Спаренные взрослые особи S. haematobium локализуются в венозных сплетениях мочевого пузыря, S. mansoni, S. intercalatum japonicum — в разветвлениях брыжеечных и воротной вен. В венулах этих сплетений происходит яйцекладка. Яйца из сосудов, в зависимости от вида шистосом, продвигаются в просвет мочевого пузыря или кишечника. Миграция гельминтов и яиц травмирует ткани, а продукты их распада и жизнедеятельности вызывают аллергические проявления, характерные для острой стадии болезни. В результате повреждения тканей яйцами в хронической стадии развивается воспалительно-пролиферативный процесс, вокруг яиц, задержавшихся в тканях, образуются гранулемы.
Внедрение в кожу церкариев иногда вызывает местную воспалительную реакцию (зуд купальщиков). Через 3—8 нед. у неиммунных лиц возникает лихорадка, зудящая сыпь, головная боль. Отмечаются бронхит с астматическим компонентом, гепатомегалия, лейкоцитоз с эозинофилией, возможны энцефалит, менингоэнцефалит. Длительность острой стадии от 2 нед. до 3 мес. Специфические проявления болезни развиваются в хронической стадии.
При мочеполовом Ш. через 3—6 мес. после заражения появляется характерный признак — терминальная гематурия без дизурических явлений. Учащенное болезненное мочеиспускание, боль в надлобковой области и в промежности возникают при развитии язв на слизистой оболочке мочевого пузыря и цистита, обусловленного вторичной инфекцией. В поздних стадиях болезнь может осложниться гидронефрозом, пиелонефритом. Поражение половых органов у женщин проявляется в виде кольпита, эрозий и полипов шейки матки, у мужчин — в виде эпидидимита, простатита, везикулита.
При кишечном шистосомозе Мансона основные проявления связаны с поражением дистального отдела толстой кишки: жидкий стул с примесь слизи и крови, чередующийся с запорами, боль в животе схваткообразного или ноющего характера, тенезмы. При заносе шистосом в легкие в них возникает воспалительный процесс, проявляющийся лихорадкой, кашлем, одышкой, болью при дыхании. При развитии гипертензии в малом круге кровообращения появляются признаки хронического легочного сердца. Японский Ш. напоминает кишечный, но протекает тяжелее. Кишечный интеркалатный Ш. индентичен шистосомозу Мансона, течение его отличается доброкачественностью; прогноз благоприятный.
Диагноз устанавливают на основании клинической картины, данных эпидемиологического анамнеза, результатов лабораторных исследований. Наиболее достоверным является обнаружение яиц в осадке мочи. В основе диагностики кишечного Ш. лежит выявление яиц шистосом Мансона в кале. Важное значение имеют также ректороманоскопия, колоноскопия. При всех формах Ш. в качестве методов предварительной диагностики можно использовать иммунологические тесты. Дифференциальный диагноз в острой стадии болезни проводят с брюшным тифом (Брюшной тиф) и сыпным тифом эпидемическим (Сыпной тиф), малярией (Малярия), туберкулезом (Туберкулёз). В хронической стадии шистосомоза Мансона и японского Ш. болезнь дифференцируют с дизентерией (Дизентерия), новообразованием толстой кишки, амебиазом (Амёбиаз). Мочеполовой Ш. дифференцируют с мочекаменной болезнью (Мочекаменная болезнь).
Лечение больных легкими формами Ш. можно проводить амбулаторно. Для специфического лечения всех Ш. используют ниридазол (амбильгар), метрифонат, гакантон (этренол), празиквантел (бильтрицид), оксамнихин.
Прогноз при своевременном распознавании и лечении благоприятный. В случаях японского Ш. прогноз серьезный в любой стадии болезни.
Профилактика включает комплекс мероприятий, направленных на раннее выявление и лечение больных, уничтожение промежуточных хозяев — моллюсков, гигиеническое воспитание населения. Для уничтожения моллюсков используют химические препараты и средства растительного происхождения, расселяют в водоемах хищников, которые уничтожают яйца, церкарии и моллюсков. Большое значение имеет санитарное благоустройство населенных мест. Специалисты, выезжающие в эндемичные регионы, должны быть предупреждены об опасности заражения при купании в пресноводных водоемах, хождении босиком по прибрежной траве и пр.
71. Ленточные черви (лентецы, цестоды) (лат. Cestoda) — класс паразитических плоских червей (Plathelminthes). Описано около 3500 видов. Представители этого таксона полностью утратили пищеварительную систему. Некоторые виды — опасные паразиты человека; вызываемые ими заболевания носят название цестодозы.
В традиционной трактовке жизненный цикл цестод представляет собой онтогенез одной особи, сменяющей нескольких хозяев. Дефинитивные (способные к половому размножению) стадии паразитируют в кишечнике водных и наземных позвоночных. Стадии, обитающие в промежуточных хозяевах (позвоночных и беспозвоночных), могут локализоваться в тканях и полостях тела.
Жизненный цикл
Жизненный цикл может состоять из 3—4 этапов. На первом этапе взрослые черви обитают в кишечнике окончательного хозяина, размножаются и продуцируют яйца. На втором этапе яйца попадают во внешнюю среду: в почву или в воду. На суше в яйцах формируется личинка или зародыш, представляющий фазу внедрения в промежуточного хозяина. У некоторых видов, яйца которых развиваются в воде, из яйца выходит свободноплавающая личинка, покрытая ресничками, а в ней формируется вторая личиночная фаза. На третьем этапе происходит развитие личинок в промежуточном хозяине, где развиваются в пузырчатую глисту — финну. Финна для дальнейшего развития должна попасть в кишечник основного хозяина, где головка из финны выворачивается, присасывается к стенке кишечника, после чего начинается процесс роста червя.
Строение
В связи с паразитизмом у большинства ленточных червей редуцирована пищеварительная система, слабо развиты нервная система и органы чувств. С другой стороны, у них сильно развита половая система, метамерно повторяющаяся в члениках, что обеспечивает их высокую плодовитость как паразитов. Это повышает возможность выживания ленточных червей, развивающихся со сменой хозяев.
Данный класс характеризуется длинным (от долей миллиметра до 10 метров) лентовидным телом. На переднем конце тела имеется «головка» — сколекс с органами прикрепления (присосками, иногда дополненными хитиновыми крючками). За головкой расположена шейка, являющаяся зоной роста ленточного червя. От шейки постоянно отшнуровываются новые членики (имеются черви и с нерасчленённым телом), последовательность которых (числом от двух до нескольких тысяч) называется стробила[1]. На заднем конце тела зрелые членики с яйцами отрываются и выносятся с фекалиями хозяина во внешнюю среду; а за счёт образования новых члеников в области шейки тело паразита восстанавливается.
Тело червей покрыто тегументом, состоящим из наружного цитоплазматического слоя клеток, которые имеют вытянутую форму, благодаря которой ядра находятся в погружённом слое. Поверхность тегумента образует микротрихии, имеющие значение при всасывании пищи из кишечника хозяина. Наружный слой тегумента подстилает мембрана, под которой располагаются кольцевые и продольные мышцы. Как и у других плоских червей, у ленточных имеются пучки дорсовентральных мышц.
Нервная система ортогональна. В сколексе имеется парный ганглий, от которого отходит несколько пар нервных тяжей. Наиболее развиты два боковых ствола. В коже червей располагаются осязательные и рецепторные клетки.
Половая система червей гермафродитная и повторяется в каждом членике. У видов с нерасчлененным телом может быть одиночный половой аппарат или метамерный ряд половых аппаратов. У цестод небольших размеров оплодотворение перекрёстное. Крупные формы (солитеры) встречаются в кишечнике хозяина поодиночке, что делает перекрёстное оплодотворение невозможным. При половом размножении у них происходит копуляция разных члеников одной особи. Самооплодотворение в пределах одного членика очень редко. Плодовитость цестод чрезвычайно велика, например, бычий солитёр в год продуцирует около 600 млн. яиц, а за всю жизнь (18—20 лет) он может производить около 11 млрд. яиц[2].
Некоторые виды при отсутствии еды могут съесть 95 % своего тела[3].
Ленточные черви или лентецы или цестоды (лат.Eucestoda)— класс паразитических плоских червей (Platyhelmithes). Цестоды — слово греческого происхождения. Корень этого слова на русский язык переводится как "пояс", "лента". Отсюда русское название класса — Ленточные черви. И действительно, форма тела животных, объединяемых данным классом, в типичных случаях очень напоминает ленту.
Описано около 3500 видов. Полностью утратили пищеварительную систему. Некоторые виды— опасные паразиты человека, вызываемые ими заболевания— цестодозы
В традиционной трактовке жизненный цикл цестод представляет собой онтогенез одной особи, сменяющей нескольких хозяев. Дефинитивные (способные к половому размножению) стадии паразитируют в кишечнике водных и наземных позвоночных. Стадии, обитающие в промежуточных хозяевах (позвоночных и беспозвоночных), могут локализоваться в тканях и полостях тела.
Все цестоды ведут паразитический образ жизни. В половозрелой стадии они обитают в теле рыб, амфибий, рептилий, птиц и млекопитающих. Личинки же их поселяются в различных беспозвоночных, главным образом членистоногих. Есть и такие виды — это наиболее опасные паразиты, личинки которых, как и взрослые особи, обитают в организме позвоночных животных.
В теле окончательных хозяев — у позвоночных животных и человека — половозрелые цестоды поселяются обычно в просвете кишечника, прикрепляясь передним концом к его внутренней стенке. Известно лишь несколько видов, приспособившихся к обитанию в желудке, в клоаке (у птиц) и в протоках печени. В этом отношении цестоды отличаются от родственного им класса трематод, которые, как отмечалось выше, приспособились к обитанию почти во всех органах своих хозяев.
72. Свиной цепень. Морфология, цикл развития, профилактика
Свиной, или вооруженный, цепень (Taenia solium) — возбудитель тениоза. Заболевание встречается повсеместно в районах, где население употребляет в пищу сырое или недостаточно термически обработанное свиное мясо.
В теле человека паразит обитает в тонком кишечнике, может быть обнаружен в глазах, центральной нервной системе, печени, мышцах, легких.
Половозрелые формы достигают в длину 2—3 м. На головке имеются присоски, а также венчик из 22—32 крючьев.
Гермафродитные проглоттиды имеют мужской половой аппарат, который состоит из нескольких сотен семенников и извилистого семяизвергательного канала, переходящего в циррусную сумку.
Она переходит в клоаку и открывается наружу. Имеются отличительные признаки в строении женской половой системы. Яичник имеет третью дополнительную дольку и большее количество ветвей (7—12), что является важным диагностическим признаком. Яйца не отличаются от яиц бычьего цепня.
Жизненный цикл. Окончательный хозяин — только человек. Промежуточные хозяева — свинья, изредка человек. Характерная особенность: членики выделяются с фекалиями человека не по одному, а группами по 5—6 штук. При подсыхании яиц их оболочка лопается, яйца свободно рассеиваются. Этому процессу также способствуют мухи и птицы.
Свиньи заражаются, поедая нечистоты, в которых могут содержаться проглоттиды. В желудке свиней растворяется оболочка яйца, из него выходят шестикрючные онкосферы. По кровеносным сосудам они попадают в мышцы, где оседают и через 2 месяца превращаются в финны. Они носят названия цистицер-ков и представляют собой пузырек, заполненный жидкостью, внутрь которого ввернута головка с присосками. В свинине цисти-церки имеют размер рисового зернышка и видны невооруженным глазом.
Заражение человека происходит при употреблении в пищу сырой или недостаточно термически обработанной свинины. Под действием пищеварительных соков оболочка цистицерки растворяется; выворачивается сколекс, который прикрепляется к стенке тонкого кишечника. Затем от шейки начинают образовываться новые проглоттиды. Через 2—3 месяца паразит достигает половой зрелости и начинает продуцировать яйца.
При этом заболевании довольно часто возникают обратная перистальтика кишечника и рвота. При этом зрелые членики попадают в желудок и перевариваются там под влиянием желудочного сока. Освободившиеся онкосферы попадают в сосуды кишечника и с током крови разносятся по органам и тканям. Могут попадать в печень, головной мозг, легкие, глаза, где формируют цистицерки. Цистицеркоз головного мозга часто является причиной смерти больных, а цистицеркоз глаз приводит к потере зрения.
Лечение цистицеркоза только хирургическое.
Диагностика
Обнаружение в фекалиях больного зрелых члеников, имеющих специфическое строение. Членики можно обнаружить и на теле и белье человека, так как они могут выползать из ануса и активно передвигаться.
Профилактика.
1.Личная. Тщательная термическая обработка свинины.
2.Общественная. Охрана пастбищ от заражения фекалиями человека. Строгий надзор за обработкой и продажей мяса на мясокомбинатах, бойнях, рынках.
Жизненный цикл свиного цепня
Два вида этих гельминтов различаются промежуточными хозяевами. Постоянным хозяином цепней является человек. Яйца этих червей - паразитов с загрязненной почвы попадают в желудки быков, коров или свиней.
Свиной цепень вызывает тениоз. В кишечнике свиньи из яйца развивается личинка, названная онкосферой, представляющая собой шарик из множества клеток с шестью хитиновыми крючками на нем.
Активно работая этими крючками, личинки проникают в кровь и переносятся в мышцы и другие органы.
Там яйца преобразуются в личиночные пузырьки — цистицерки (финны). Это особая стадия развития всех ленточных червей, ее еще называют пузырчатой глистой. Внешне финна свиного цепня выглядит как пузырек размером с горошину, наполненный жидкостью.
При внимательном рассмотрении можно заметить на финне небольшую ямку, внутри которой спрятаны неразвившиеся присоски и крючья. Это будущая головка червя (сколекс), втянутая внутрь финны.
В этот момент рост и развитие финны приостанавливаются. Для дальнейшего преобразования финне необходимо попасть в кишечник человека [3,c.49].
Если мясо зараженного животного не подвергнется достаточной тепловой обработке и попадет в организм человека, личинки активизируются, и это станет началом развития взрослой особи гельминта.
В кишечнике человека финна выворачивает наружу втянутую до этого головку. Присосками и крючьями она крепко цепляется к стенкам кишечника, и солитер начинает наращивать от головки новые членики.
Взрослая особь свиного цепня представляет собой червя с лентовидным телом белого цвета, размерами от 1,5 до 6 м. Головка цепня микроскопических размеров (2-3 мм), имеет венчик крючьев и 4 присоски. За головкой расположена шейка, а за ней — членики, имеющие квадратную форму.
От свиного цепня легче избавиться, чем от бычьего, но он гораздо опаснее, потому что может паразитировать не только в кишечнике человека.
Финны этого гельминта могут находиться в мозге, печени и даже в глазах. Подобные случаи паразитирования цепня неизлечимы и часто заканчиваются смертью большого.
Заражение происходит через грязные руки, несвежее белье, продукты. Для человека, в кишечнике которого паразитирует свиной солитер, существует опасность, что при рвоте личинки гельминта могут попасть из кишечника в желудок, откуда онкосферы разнесут финны по другим органам.
Симптомы заболевания бывают самыми разными: тошнота, рвота, понос, отсутствие аппетита. При диагностике недостаточно обнаружить яйца гельминта в фекалиях, так как онкосферы бычьего и свиного цепней нельзя отличить друг от друга. Необходимо обнаружить и исследовать зрелые членики[6,c.41].
Человек может служить и промежуточным хозяином для свиного цепня, когда заражение происходит от другого человека. Цистицерки, образовавшиеся в органах человека, вызывают цистицеркоз, который может проявляться в судорожных припадках и слепоте.
Способные к размножению членики имеют длину 1,6-3 см и ширину 5-7 мм. Зрелые членики начинаются с двухсотой проглоттиды и далее. Они могут сами выползать из анального отверстия хозяина, передвигаться по телу и белью.
95. История становления эволюционных идей. Сущность представлений Ч. Дарвина о механизмах эволюции живой природы. Синтетическая теория эволюции.
Биологическая эволюция - это необратимое и направленное историческое развитие живой природы, сопровождающееся изменением генетического состава популяций, формированием адаптации, образованием и вымиранием видов, преобразованиями биогеоценозов и биосферы в целом. Иными словами, под биологической эволюцией следует понимать процесс приспособительного исторического развития живых форм на всех уровнях организации живого.
В додарвиновский период (до 1859 г.) в естествознании господствовали метафизические взгляды на природу, когда явления и тела природы рассматривались как раз и навсегда данные, неизменные, изолированные, не связанные между собой. Они были тесно связаны с креационизмом и теологией (греч. teos — боги), согласно которым многообразие органического мира есть результат творения его Богом. Креационисты (К. Линней, Ж. Кювье) доказывали, что виды живой природы реальны и неизменны со времени своего появления, при этом К. Линней утверждал, что видов существует столько, сколько их было создано во время «сотворения мира».
К концу XVIII в. в биологии накопился огромный описательный материал, который позволял сделать следующие выводы: 1) даже внешне далекие виды по внутреннему строению обнаруживают определенные черты сходства; 2) современные виды отличаются от давно живших на Земле ископаемых; 3) внешний вид, строение и продуктивность сельскохозяйственных растений и животных могут существенно изменяться в зависимости от условий их выращивания и содержания.
Появившиеся сомнения в неизменности видов привели к возникновению трансформизма — системы взглядов об изменяемости и превращении форм растений и животных под влиянием естественных причин.
Идеи трансформизма нашли дальнейшее развитие в трудах выдающегося французского биолога Ж. Б. Ламарка (1744— 1829) — создателя первого эволюционного учения.
Занимаясь классификацией животных, Ламарк пришел к выводу, что виды не остаются постоянными, они медленно и непрерывно изменяются. В его системе, в отличие от системы Линнея, животные размешены в восходящем порядке — от инфузорий и полипов до высокоорганизованных существ (птицы и млекопитающие). Ламарк впервые в истории биологии сформулировал положение об эволюционном развитии живой природы: жизнь возникает путем самозарождения простейших живых тел из веществ неживой природы. Дальнейшее развитие идет по пути прогрессивного усложнения организмов, т. е. путем эволюции. В попытке найти движущие силы прогрессивной эволюции Ламарк пришел к произвольному выводу, что в природе существует некий изначальный закон внутреннего стремления организмов к совершенствованию. Согласно этим представлениям, все живое, начиная с самозародившихся инфузорий, постоянно стремится к усложнению своей организации в длинном ряду поколений, что в конечном итоге приводит к превращению одних форм живых существ в другие (например, инфузории постепенно превращаются в полипов, полипы — в лучистых и т. д.). Главным фактором изменчивости организмов Ламарк считал влияние внешней среды: изменяются условия (климат, пища), а вслед за этим из поколения в поколение изменяются и виды. Таким образом, изменения органов, возникающие как прямым, так и косвенным путем, становятся, по Ламарку, сразу полезными, приспособительными. Если изменения организмов, вызванные прямым или косвенным воздействием условий среды, повторяются в целом ряду поколений, то они наследуются и становятся признаками новых видов. Ламарковское толкование причин изменения видов в природе имеет серьезные недостатки. Так, влиянием упражнения или неупражнения органов нельзя объяснить изменения таких признаков, как длина волосяного покрова, густота шерсти, жирность молока, окраска покровов животных, которые не могут упражняться. Кроме того, как теперь известно, не все изменения, возникающие у организмов под влиянием окружающей среды, наследуются.
После Ламарка появляется теория Дарвина. Теория Дарвина представляет собой целостное учение об историческом развитии органического мира. Сущность эволюционного учения заключается в следующих основных положениях:
1) Все виды живых существ, населяющих Землю, никогда не были кем-то созданы.
2) Возникнув естественным путем, органические формы медленно и постепенно преобразовывались и совершенствовались в соответствии с окружающими условиями.
3) В основе преобразования видов в природе лежат такие свойства организмов, как изменчивость и наследственность, а также постоянно происходящий в природе естественный отбор. Естественный отбор осуществляется через сложное взаимодействие организмов друг с другом и с факторами неживой природы; эти взаимоотношения Дарвин назвал борьбой за существование.
4) Результатом эволюции является приспособленность организмов к условиям их обитания и многообразие видов в природе.
Синтетическая теория эволюции — современный дарвинизм — возникла в начале 40-х годов XX в. Она представляет собой учение об эволюции органического мира, разработанное на основе данных современной генетики, экологии и классического дарвинизма.
Основные положения синтетической теории эволюции:
Материалом для эволюции служат наследственные изменения — мутации (как правило, генные) и их комбинации.
Основным движущим фактором эволюции является естественный отбор, возникающий на основе борьбы за существование.
Наименьшей единицей эволюции является популяция.
Эволюция носит в большинстве случаев дивергентный характер, т. е. один таксон может стать предком нескольких дочерних таксонов.
Эволюция носит постепенный и длительный характер. Видообразование как этап эволюционного процесса представляет собой последовательную смену одной временной популяции чередой последующих временных популяций.
Вид состоит из множества соподчиненных, морфологически, физиологически, экологически, биохимически и генетически отличных, но репродуктивно не изолированных единиц — подвидов и популяций.
Вид существует как целостное и замкнутое образование. Целостность вида поддерживается миграциями особей из одной популяции в другую, при которых наблюдается обмен аллелями («поток генов»),
Макроэволюция на более высоком уровне, чем вид (род, семейство, отряд, класс и др.), идет путем микроэволюции. Согласно синтетической теории эволюции, не существует закономерностей макроэволюции, отличных от микроэволюции. Иными словами, для эволюции групп видов живых организмов характерны те же предпосылки и движущие силы, что и для микроэволюции.
Любой реальный (а не сборный) таксон имеет монофилети-ческое происхождение.
Эволюция имеет ненаправленный характер, т. е. не идет в направлении какой-либо конечной цели.
96. Происхождение жизни: гипотезы панспермии и абиогенного происхождения жизни. Главные этапы возникновения и развития жизни.
Существуют две главные гипотезы, по-разному объясняющие появление жизни на Земле. Согласно гипотезе панспермии, жизнь занесена из космоса либо в виде спор микроорганизмов, либо путем намеренного «заселения» планеты разумными пришельцами из других миров.
Согласно второй гипотезе, жизнь возникла на Земле, когда сложилась благоприятная совокупность физических и химических условий, сделавших возможным абиогенное образование органических веществ из неорганических.
Главные этапы на пути возникновения и развития жизни, по-видимому, состоят в: 1) образовании атмосферы из газов, которые могли бы служить «сырьем» для синтеза органических веществ (метана, оксида и диоксида углерода, аммиака, сероводорода, цианистых соединений), и паров воды; 2) абиогенном (т.е. происходящем без участия организмов) образовании простых органических веществ, в том числе мономеров биологических полимеров — аминокислот, сахаров, азотистых оснований, АТФ и других мононуклеотидов; 3) полимеризации мономеров в биологические полимеры, прежде всего белки (полипептиды) и нуклеиновые кислоты (полинуклеотиды); 4) образовании предбиологических форм сложного химического состава — протобионтов, имеющих некоторые свойства живых существ; 5) возникновении простейших живых форм, имеющих всю совокупность главных свойств жизни,—примитивных клеток; 6) биологической эволюции возникших живых существ.
Это можно не говорить, но для общего развития пойдет!!! Это предыдущий абзац – полная версия. Возможность абиогенного образования органических веществ, включая мономеры биологических полимеров, в условиях, бывших на Земле около 4 млрд. лет назад, доказана опытами химиков. В лабораторных условиях при пропускании электрических разрядов через различные газовые смеси, напоминающие примитивную атмосферу планеты, а также при использовании других источников энергии ученые получали среди продуктов реакций аминокислоты (аланин, глицин, аспарагиновую кислоту), янтарную, уксусную, молочную кислоты, мочевину, азотистые основания (аденин, гуанин), АДФ и АТФ. Низкомолекулярные органические соединения накапливались в водах первичного океана в виде первичного бульона или же адсорбировались на Поверхности глинистых отложений. Последнее повышало концентрацию этих веществ, создавая тем самым лучшие условия для полимеризации.
Возможность полимеризации низкомолекулярных соединений с образованием полипептидов и полинуклеотидов (определяющая следующий этап на пути возникновения жизни) непосредственно в первичном бульоне вызывает сомнения по термодинамическим соображениям. Водная среда благоприятствует реакции деполимеризации. Ученые предполагают, что образование полипептидов и полинуклеотидов могло происходить в пленке из низкомолекулярных органических соединений в безводной среде, например на склонах вулканических конусов, покрытых остывающей лавой. Это предположение находит подтверждение в опытах. Выдерживание в течение определенного времени при 130°С сухой смеси аминокислот в сосудах из кусков лавы приводило к образованию полипептидов.
Образующиеся описанным образом биополимеры смывались ливневыми потоками в первичный бульон, что защищало их от разрушающего действия УФ-излучения, которое в то время из-за отсутствия в атмосфере планеты озонового слоя было очень жестким.
По мере повышения концентрации полипептидов, полинуклеотидов и других органических соединений в первичном бульоне сложились условия для следующего этапа — самопроизвольного возникновения предбиологических форм сложного химического состава, или протобионтов. Предположительно они могли быть представлены коацерватами (А. И. Опарин) или микросферами (С. Фоке). Это коллоидные капли с уплотненным поверхностным слоем, имитирующим мембрану, содержимое которых составляли один или несколько видов биополимеров. Возможность образования в коллоидных растворах структур типа коацерватов или микросфер доказана опытным путем.
При определенных условиях коацерваты проявляют некоторые общие свойства живых форм. Они способны до известной степени избирательно поглощать вещества из окружающего раствора. Часть продуктов химических реакций, проходящих в коацерватах с участием поглощаемых веществ, выделяется ими обратно в среду. Происходит процесс, напоминающий обмен веществ. Накапливая вещества, коацерваты увеличивают свой объем (рост). По достижении определенных размеров они распадаются на части, сохраняя при этом некоторые черты исходной химической организации (размножение). Поскольку устойчивость коацерватов различного химического состава различна, среди них происходит отбор.
Перечисленные выше свойства ученые усматривают у протобионтов. Протобионты представляются как обособленные от окружающей среды, открытые макромолекулярные системы, возникавшие в первичном бульоне и способные к примитивным формам роста, размножения, обмена веществ и предбиологическому химическому отбору.
Предбиологическая эволюция протобионтов осуществлялась в трех главных направлениях. Важное значение имело совершенствование каталитической (ферментной) функции белков. Один из путей, дающих требуемый результат, заключается, по-видимому, в образовании комплексов металлов с органическими молекулами. Так, включение железа в порфириновое кольцо Гемоглобина увеличивает его каталитическую активность в сравнении с активностью самого железа в растворе в 1000 раз. Развивалось такое свойство биологического катализа, как специфичность. Во-вторых, исключительная роль в эволюции протобионтов принадлежит приобретению полинуклеотидами способности к самовоспроизведению, что сделало возможным передачу информации от поколения к поколению, т.е. сохранение ее во времени. В основе этой способности лежит матричный синтез. Механизм матричного синтеза был использован также для переноса информации с полинуклеотидов на полипептиды. Третье главное направление эволюции протобионтов состояло в возникновении мембран. Отграничение от окружающей среды мембраной с избирательной проницаемостью превращает протобионт в устойчивый набор макромолекул, стабилизирует важные параметры обмена веществ на основе специфического катализа.
Разделение функций хранения и пространственно-временной передачи информации, с одной стороны (нуклеиновые кислоты), и использование ее для организации специфических структуры и обмена веществ — с другой (белки); появление молекулярного механизма матричного синтеза биополимеров; освоение эффективных систем энергообеспечения жизнедеятельности (АТФ); образование типичной биологической мембраны — все это привело к возникновению живых существ, которые поначалу были представлены примитивными клетками.
С момента появления клеток предбиологический химический отбор уступил место биологическому отбору. Дальнейшее развитие жизни шло согласно законам биологической эволюции. Переломным моментом на этом пути было возникновение клеток эукариотического типа, многоклеточных организмов, человека.
97. Появление клетки как исходная точка биологической эволюции. Гипотезы происхождения эукариотических клеток (симбиотическая, инвагинационная). Возникновение многоклеточности. Отличия клеток одно- и многоклеточных организмов.
Ископаемые останки клеток эукариотического типа обнаружены в породах, возраст которых не превышает 1,0—1,4 млрд. лет. Эукариотические клетки произошли от предка, имевшего прокариотическое строение.
Наиболее популярна в настоящее время симбиотическая гипотеза происхождения эукариотических клеток, согласно которой основой, или клеткой-хозяином, в эволюции клетки эукариотического типа послужил анаэробный прокариот, способный лишь к амебоидному движению. Переход к аэробному дыханию связан с наличием в клетке митохондрии, которые произошли путем изменений симбионтов — аэробных бактерий, проникших в клетку-хозяина и сосуществовавших с ней.
Сходное происхождение предполагают для жгутиков, предками которых служили симбионты-бактерии, имевшие жгутик и напоминавшие современных спирохет. Приобретение клеткой жгутиков имело наряду с освоением активного способа движения важное следствие общего порядка. Предполагают, что базальные тельца, которыми снабжены жгутики, могли эволюционировать в центриоли в процессе возникновения механизма митоза.
Способность зеленых растений к фотосинтезу обусловлена присутствием в их клетках хлоропластов. Сторонники симбиотической гипотезы считают, что симбионтами клетки-хозяина, давшими начало хлоропластам, послужили прокариотические синезеленые водоросли.
Серьезным доводом в пользу симбиотического происхождения митохондрий, центриолей и хлоропластов является то, что перечисленные органеллы имеют собственную ДНК. Вместе с тем белки бациллин и тубулин, из которых состоят жгутики и реснички соответственно современных прокариот и эукариот, имеют различное строение.
Внутриклеточные мембраны гладкой и шероховатой цитоплазматической сети, пластинчатого комплекса, пузырьков и вакуолей рассматривают как производные наружной мембраны ядерной оболочки, которая способна образовывать впячивания.
Центральным и трудным для ответа является вопрос о происхождении ядра. Предполагают, что оно также могло образоваться из симбионта-прокариота. Увеличение количества ядерной ДНК, во много раз превышающее в современной эукариотической клетке ее количество в митохондрий или хлоропласте, происходило, по-видимому, постепенно путем перемещения групп генов из геномов симбионтов. Нельзя исключить, однако, что ядерный геном формировался путем наращивания генома клетки-хозяина (без участия симбионтов).
Согласно инвагинационной гипотезе, предковой формой эукариотической клетки был аэробный прокариот. Внутри такой клетки-хозяина находилось одновременно несколько геномов, первоначально прикреплявшихся к клеточной оболочке. Органеллы, имеющие ДНК, а также ядро, возникли путем впячивания и отшнуровывания участков оболочки с последующей функциональной специализацией в ядро, митохондрий, хлоропласты. В процессе дальнейшей эволюции произошло усложнение ядерного генома, появилась система цитоплазматических мембран.
Большое значение при переходе к многоклеточности имело наличие у эукариотических клеток эластичной оболочки, что необходимо для образования устойчивых клеточных комплексов.
Среди цитофизиологических особенностей эукариот, увеличивающих их эволюционные возможности, необходимо назвать аэробное дыхание, которое также послужило предпосылкой для развития многоклеточных форм. Интересно, что сами эукариотические клетки появились на Земле после того, как концентрация O2 в атмосфере достигла 1% (точка Пастера). Названная концентрация является необходимым условием аэробного дыхания.
Закономерность, проявляющаяся в разделении и специализации функций и структур, представляет собой одно из всеобщих свойств жизни. Возникновение среди живых форм многоклеточных организмов, с которыми связано прогрессивное направление эволюции, является логичным развитием этого свойства. В таких организмах усиление жизненной силы благодаря многократному повторению клеточных механизмов сочетается с широчайшим размахом разделения функций, их совершенствованием, образованием разнообразных специализированных структур — органов и их систем.
Переход к многоклеточности — одновременно и новое качественное состояние жизни, для которого характерно ускорение эволюционных преобразований на основе более полного использования резерва наследственной изменчивости. Это обусловлено, во-первых, объединением у многоклеточных организмов полового процесса и размножения в единое целое — половое размножение. Во-вторых, хотя цикл индивидуального развития имеют все живые формы, включая вирусы, только у многоклеточных организмов выделяется эмбриональный период. Отмеченные особенности многоклеточной организации живых существ сделали их основой дальнейшей прогрессивной эволюции. Эволюционными предшественниками многоклеточных организмов были колониальные формы простейших организмов. Наиболее ранние ископаемые останки многоклеточных животных имеют возраст около 700 млн. лет.
Многоклеточные организмы отличаются от колониальных прежде всего разделением функций отдельных групп клеток при взаимодействии со средой. Принципиальное отличие проявляется в процессе самовоспроизведения клетки. Дочерняя клетка не всегда становится полной копией материнской клетки. На процесс самовоспроизведения клетки оказывает влияние ее информационное взаимодействие с окружающими клетками и внешней средой организма. ДНК в дочерней клетке полностью копируется с ДНК материнской клетки, а комплекс остальных элементов может значительно отличаться.
Благодаря отмеченным особенностям за 1 млрд. лет эволюции эукариотический тип клеточной организации дал широкое разнообразие живых форм от одноклеточных простейших до млекопитающих и человека.
98. Элементарные эволюционные факторы: мутационный процесс, генетическая комбинаторика, популяционные волны и генетико-автоматические процессы.
События и процессы, способствующие преодолению генетической инертности популяций и приводящие к изменению их генофондов, называют элементарными эволюционными факторами. Важнейшими из них являются мутационный процесс, популяционные волны, изоляция, естественный отбор.
Мутационный процесс - это спонтанный процесс возникновения мутаций. Мутационный процесс, выполняя роль элементарного эволюционного фактора, происходит постоянно на протяжении всего периода существования жизни, а отдельные мутации возникают многократно у разных организмов. Генофонды популяций испытывают непрерывное давление мутационного процесса. Это обеспечивает накопление мутаций, несмотря на высокую вероятность потери в ряду поколений единичной мутации. Благодаря этому процессу достигается тройственный результат: 1) исключается непосредственное отрицательное влияние мутантного аллеля на фенотипическое выражение признака, контролируемого данным геном; 2) сохраняются нейтральные мутации, не имеющие приспособительной ценности в настоящих условиях существования, но которые смогут приобрести такую ценность в будущем; 3) накапливаются некоторые неблагоприятные мутации, которые в гетерозиготном состоянии нередко повышают относительную жизнеспособность организмов (эффект гетерозиса). Таким образом создается резерв наследственной изменчивости популяции. Как правило, один фенотипический признак формируется под влиянием множества генетических факторов, то есть является полигенным. Поэтому разные мутации могут взаимодействовать между собой. Поэтому генетическая комбинаторика выделяется как отдельный эволюционный фактор.
Популяционными волнами или волнами жизни называют периодические или апериодические колебания численности организмов в природных популяциях. Причины колебаний часто имеют экологическую природу. Изменение генофондов популяций происходит как на подъеме, так и на спаде популяционной волны. При росте численности организмов наблюдается слияние ранее разобщенных популяций и объединение их генофондов. Так как популяции по своему генетическому составу уникальны, в результате такого слияния возникают новые генофонды с измененными по сравнению с исходными частотами аллелей. В условиях возросшей численности интенсифицируются межпопуляционные миграции особей, что также способствует перераспределению аллелей. Рост количества организмов обычно сопровождается расширением занимаемой территории. На гребне популяционной волны некоторые группы особей выселяются за пределы ареала вида и оказываются в необычных условиях существования. В таком случае они испытывают действие новых факторов естественного отбора. Повышение концентрации особей в связи с ростом их численности усиливает внутривидовую борьбу за существование. При спаде численности наблюдается распад крупных популяций. Возникающие малочисленные популяции характеризуются измененными генофондами. В условиях массовой гибели организмов редкие мутантные аллели могут быть генофондом потеряны. При сохранении редкого аллеля его концентрация в генофонде малочисленной популяции автоматически возрастает.
Случайные, но не обусловленные действием естественного отбора колебания частот аллелей называют генетико-автоматическими процессами или дрейфом генов. В результате происходят гомозиготизация особей и затухание изменчивости. Дрейф генов обусловливает утрату или закрепление аллелей в гомозиготном состоянии у всех членов популяции вне связи с их приспособительной ценностью. Он играет важную роль в формировании генофондов малочисленных групп организмов, изолированных от остальной части вида.
99. Естественный отбор – движущая и направляющая сила эволюции. Формы естественного отбора.
В природных популяциях организмов, размножающихся половым способом, существует большое разнообразие генотипов и, следовательно, фенотипов. Различия между организмами по приспособленности, оцениваемой передачей аллелей следующему поколению, выявляются в природе с помощью естественного отбора. Естественный отбор — процесс, изначально определённый Чарльзом Дарвином как приводящий к выживанию и преимущественному размножению более приспособленных к данным условиям среды особей, обладающих полезными наследственными признаками. Главный результат отбора заключается не просто в выживании более жизнеспособных, а в относительном вкладе таких особей в генофонд дочерней популяции. Необходимой предпосылкой отбора является борьба за существование – конкуренция за пищу, жизненное пространство, партнера для спаривания. Естественный отбор происходит на всех стадиях онтогенеза организмов. Благодаря отбору аллели (признаки), повышающие выживаемость и репродуктивную способность, накапливаются в ряду поколений, изменяя генетический состав популяций в биологически целесообразном направлении.
В природных условиях естественный отбор осуществляется исключительно по фенотипу. Отбор генотипов происходит вторично через отбор фенотипов, которые отражают генетическую конструкцию организмов.
Как элементарный эволюционный фактор естественный отбор действует в популяциях. Популяция является полем действия, отдельные особи – объектами действия, а конкретные признаки – точками приложения отбора.
Отбор, сохраняющий определенные фенотипы, по своему направлению является положительным, тогда как отбор, устраняющий фенотипы из популяции, - отрицательным.
В зависимости от результата различают:
Стабилизирующий отбор. Он сохраняет в популяции средний вариант фенотипа или признака. Приводит к преимущественному размножению типичных организмов. Благодаря этой форме сохраняются результаты предшествующих этапов эволюции.
Движущий отбор обуславливает последовательное изменение фенотипа в определенном направлении, что проявляется в сдвиге средних значений отбираемых признаков в сторону их усиления или ослабления. При смене условий обитания благодаря этой форме отбора в популяции закрепляется фенотип, более соответствующий среде.
Дизруптивный (разрывающий) отбор сохраняет несколько разных фенотипов с равной приспособленностью. Он действует против особей со средним или промежуточным значением признаков.
В зависимости от формы отбор сокращает масштабы изменчивости, создает новую или сохраняет прежнюю картину разнообразия. Как и другие элементарные эволюционные факторы, естественный отбор вызывает изменения соотношений аллелей в генофондах популяций. Особенность его действия состоит в том, что эти изменения направлены. Отбор приводит генофонды в соответствие с критерием приспособленности. Он осуществляет обратную связь между изменениями генофонда и условиями обитания, накладывает на эти изменения печать биологической целесообразности (полезности).
100. Вид – результат микроэволюции. Определение, структура и критерии вида. Генетическое единство, целостность вида.
Видом называют совокупность особей, сходных по основным морфологическим и функциональным признакам, кариотипу, поведенческим реакциям, имеющих общее происхождение, заселяющих определенную территорию (ареал), в природных условиях скрещивающихся исключительно между собой и при этом производящих плодовитое потомство.
Признаки (критерии) вида:
Морфологический критерий позволяет различать разные виды по внешним и внутренним признакам.
Физиолого-биохимический критерий фиксирует неодинаковость химических свойств и физиологических процессов разных видов.
Географический критерий свидетельствует, что каждый вид обладает своим ареалом.
Экологический позволяет различать виды по комплексу абиотических и биологических условий, в которых они сформировались, приспособились к жизни.
Репродуктивный критерий обуславливает репродуктивную изоляцию вида от других, даже близкородственных.
В связи с неодинаковыми условиями среды особи одного вида в пределах ареала распадаются на более мелкие единицы — популяции. Реально вид существует именно в виде популяций.
Половой процесс объединяет организмы в вид. В результате достигаются нивелировка различий между организмами внутри вида. Благодаря половому процессу происходит также объединение генов, и формируется общий генофонд. Поэтому каждый вид представляет собой генетически замкнутую систему.
101. Основные характеристики популяции как эколого-генетической системы: популяционный ареал, численность особей и ее динамика, половая и возрастная структуры, морфологическое и экологическое единство. Генофонд природных популяций, частота аллелей и генотипов. Закон Харди-Вайнберга.
Популяция с точки зрения экологии – это совокупность особей одного вида, которая имеет три критерия:
Относительная изолированность от других групп вида
Длительной проживание на одной территории
Свободное скрещивание между собой и появление плодовитого потомства.
Популяция с точки зрения генетики – это репродуктивное сообщество, обладающее общим генофондом.
Основные характеристики популяции:
Популяционный ареал (размеры ареала) – зависит от радиуса индивидуальной активности организмов данного вида и особенностей природных условий на соответствующей территории. Чем активнее животное, тем больше ареал. Например, ареал виноградной улитки меньше, чем ареал писца, так как ее активность очень мала – несколько десятков метров, а песца – несколько сотен километров.
Численность особей. Существуют минимальные значения численности, при которых популяция способна поддерживать себя во времени. Сокращение численности ниже этого минимума приводит к вымиранию популяции. Численность может иметь сезонные колебания.
Возрастная структура варьирует в зависимости от продолжительности жизни, интенсивности размножения, возраста достижения половой зрелости. В зависимости от вида организмов она может быть то более, то менее сложной.
Половой состав обусловливается эволюционно закрепленными механизмами формирования первичного (на момент зачатия), вторичного (на момент рождения) и третичного (во взрослом состоянии) соотношения полов. В качестве примера рассмотрим изменение полового состава популяции людей. На момент рождения оно составляет 106 мальчиков на 100 девочек, в возрасте 16—18 лет выравнивается, в возрасте 50 лет насчитывает 85 мужчин на 100 женщин, а в возрасте 80 лет — 50 мужчин на 100 женщин.
Популяции присуще генетическое единство. Оно обусловливается достаточным уровнем панмиксии. В условиях случайного подбора скрещивающихся особей источником аллелей для генотипов организмов последовательных поколений является весь генофонд популяции. Генетическое единство проявляется также в общей генотипической изменчивости популяции при изменении условий существования, что обусловливает как выживание вида, так и образование новых видов.
Основной закон генетики популяций сформулировали Харди и Вайнберг: «В популяции из бесконечно большого числа свободно скрещивающихся особей в отсутствие мутаций, избирательной миграции организмов с различными генотипами и давления естественного отбора первоначальные частоты аллелей сохраняются из поколения в поколение».
Уравнение частот аллелей: р+q=1
р – частота А
q – частота а
Бином Ньютона р2+2рq+q2=1, где
р2 = АА
2рq = Аа
q2 = аа
+ Доказательство (задача про двух гетерозигот)
102. Макроэволюция. Направления эволюции: ароморфоз, идиоадаптация, морфофизиологический регресс. Формы эволюции групп: филетическая, дивергентная, конвергентная, параллельная. Пути эволюции: биологический прогресс и биологический регресс.
Процесс формирования таксонов надвидового ранга называют макроэволюцией. Процесс макроэволюции изучают методами палеонтологии, сравнительной анатомии и сравнительной эмбриологии.
В зависимости от того, изменяется ли уровень организации в эволюционирующих группах, выделяют два основных типа эволюции: аллогенез и арогенез. При аллогенезе у всех представителей данной группы сохраняются без изменения основные черты строения и функционирования систем органов, благодаря чему уровень организации их остается прежним. Аллогенная эволюция происходит в пределах одной адаптивной зоны — совокупности экологических ниш, различающихся в деталях, но сходных по общему направлению действия основных средовых факторов на организм данного типа. Интенсивное заселение конкретной адаптивной зоны достигается благодаря возникновению у организмов идиоадаптаций — локальных морфофизиологических приспособлений к определенным условиям существования. Пример, особенности строения конечностей (у крота, копытных, ластоногих), особенности клюва (у хищных, куликов, попугаев).
Арогенез — такое направление эволюции, при котором у некоторых групп внутри более крупного таксона появляются новые морфофизиологические особенности, приводящие к повышению уровня их организации. Эти новые прогрессивные черты организации называют ароморфозами. Ароморфозы позволяют организмам заселять принципиально новые, более сложные адаптивные зоны. Пример, возникновение теплокровности, двух кругов кровообращения. Если в ходе филогенеза организмы осваивают более простую по сравнению с исходной адаптивную зону, например, переходя к прикрепленному неподвижному образу жизни или становись паразитами, то они претерпевают морфофизиологический регресс, или дегенерацию, утрачивая часть прогрессивных черт, которыми обладали их предки, и оказываясь на более низком уровне организации.
Существуют две элементарные формы филогенеза: филетическая и дивергентная эволюция. Филетическая эволюция — это изменения, происходящие в одном филогенетическом стволе, эволюционирующем во времени как единое целое. Реконструированы филогенетические ряды слона, лошади и других организмов, в которых можно наблюдать постепенное нарастание количества и степени выраженности признаков, характерных для современных форм.
Дивергентная эволюция заключается в образовании на основе одной предковой группы двух или нескольких производных. Она приводит к дифференциации более крупных таксонов на более мелкие. Как филетическая, так и дивергентная эволюция протекают на общей генетической базе, поэтому между организмами сохраняется более или менее выраженное генотипическое и морфофункциональное сходство.
При конвергенции генетическое родство отсутствует, однако некоторые группы оказываются вынужденными решать сходные экологические задачи, обитая в одной и той же среде. В связи с этим возникают одинаковые адаптации, затрагивающие несколько признаков. Например, сходное строение конечностей крота и медведки.
Другая форма эволюции — параллелизм — реализуется в двух или нескольких группах, связанных более или менее отдаленным родством, которое основано на дивергенции от общего предка. Например, одно- и двугорбный верблюд имеют сложные адаптации в жизни в пустынях.
Успех группы организмов в эволюционном процессе оценивают как состояние биологического прогресса, критериями которого являются: 1) увеличение количества представителей соответствующей группы; 2) расширение ареала распространения; 3) активизация видоообразования в роде, увеличение количества родов в семействе, семейств в отряде и т.д. Явление, противоположное биологическому прогрессу, — биологический регресс — свидетельствует об угасании филогенетической группы, ведущем к ее вымиранию.
103. Положение человека в системе животного мира (обоснование систематического положения вида Homo sapiens). Биологические предпосылки прогрессивного развития гоминид (антропоморфозы).
Царство: клеточные
Надцарство: эукариоты
Царство: животные
гетеротрофное питание, то есть готовыми органическими веществами
подвижный образ жизни
типичное строение клеток
Подцарство: многоклеточные
Тип: хордовые
двусторонняя симметрия
в процессе развития присутствует хорда и жаберные щели в области легких
нервная система по типу спинной нервной трубки
Подтип: позвоночные
формирование позвоночника
наличие черепа
развитый головной мозг
Класс: млекопитающие
теплокровность
наличие молочных желез
наличие волос на поверхности тела
гетеродонтная зубная система
наличие двух смен зубов
5 Отделов позвоночника
4-камернрое сердце
Одна левая дуга аорты
Подкласс: настоящие звери
живорождение
молочные железы гроздевидные
наличие сосков с протоками молочных желез
отсутствие клоаки, то есть пищеварительный и мочеполовой тракт открываются собственным отверстием.
Инфракласс: плацентарные (высшие звери)
детеныш внутри органзма матери
плод питается через плаценту матери
Отряд: приматы
конечности хватательного типа
на пальцах ногти
одна пара сосков
хорошо развиты ключицы
зубная система не специализирована, имеются все типы зубов
глаза направлены вперед
отсутствует сезонность половой жизни
рождается, как правило, один детеныш
выражена забота о потомстве
развитие мимики и звуковой сигнализации
стадные отношения
Подотряд: человекоподобные высшие приматы
ногти плоские
хорошее развитие головного мозга, особенно больших полушарий
редукция обонятельных путей
кольца трахеи не замкнутые
редукция хвостовых позвонков
хорошая способность к обучению и памяти
Семейство: гоминиды
прямохождение
кисть, приспособленная к изготовлению орудий
высокоразвитый мозг
Род: человек (Ноmо)
Вид: Человек разумный (Ноmo sapiens)
Антроморфозы – необходимые преадаптации к возникновению человека при появлении комплекса благоприятных условий. К ним относятся жизнь небольшими стадами со сложной системой иерархии и общественным воспитанием потомства, низкая плодовитость, инстинкт подражания, передача социального опыта в ряду поколений.
106. Соотношение онто- и филогенеза. Закон зародышевого сходства К. Бэра. Основной биогенетический закон Ф. Мюллера и Э. Геккеля. Понятие о рекапитуляциях и ценогенезах. Учение А.Н. Северцова о филэмбриогенезах. Провизорные и дефинитивные, гомологичные и аналогичные органы.
Сопоставляя стадии развития зародышей разных видов и классов хордовых, К. Бэр сделал следующие выводы:
Эмбрионы животных одного типа на ранних стадиях развития схожи
Они последовательно переходят в своем развитии от более общих признаков типа ко все более частным.
Эмбрионы разных представителей одного типа постепенно обособляются друг от друга
Сопоставляя онтогенез ракообразных с морфологией их вымерших предков, Ф. Мюллер сделал вывод о том, что ныне живущие ракообразные в своем развитии повторяют путь, пройденный их предками. Преобразование онтогенеза в эволюции, по мнению Ф. Мюллера, осуществляется благодаря его удлинению за счет добавления к нему дополнительных стадий или надставок. На основе этих наблюдений, а также изучения развития хордовых Э. Геккель (1866) сформулировал основной биогенетический закон, в соответствии с которым онтогенез представляет собой краткое и быстрое повторение филогенеза.
Повторение структур, характерных для предков, в эмбриогенезе потомков названо рекапитуляциями. Генетическая основа рекапитуляции заключена в единстве механизмов генетического контроля развития, сохраняющемся на базе общих генов регуляции онтогенеза, которые достаются родственным группам организмов от общих предков.
Но повторение не вызовет появления нового, а значит не приведет к эволюции поэтому необходимы изменения. Эти изменения сводятся к тому, что конкретные онтогенезы откланяются от пути, проложенного предковыми формами, и приобретают новые черты.
К таким отклонениям относятся ценогенезы — приспособления, возникающие у зародышей или личинок и адаптирующие их к особенностям среды обитания. Например, к ценогенезам у млекопитающих и человека относят зародышевые оболочки, желточный мешок, аллантоис и плаценту с пуповиной. Ценогенезы обеспечивают более высокую вероятность выживания потомства.
Другой тип филогенетически значимых преобразований филогенеза — филэмбриогенезы. Они представляют собой отклонения от онтогенеза, характерного для предков, проявляющиеся в эмбриогенезе, но имеющие адаптивное значение у взрослых форм. Например, закладки волосяного покрова появляются у млекопитающих на очень ранних стадиях эмбрионального развития, но сам волосяной покров имеет значение только у взрослых организмов.
В зависимости от того, на каких этапах эмбриогенеза возникают изменения развития, имеющие значение филэмбриогенезов, различают три их типа.
1. Анаболии возникают после того, как орган практически завершил свое развитие, и выражаются в добавлении дополнительных стадий, изменяющих конечный результат. Например, приобретение специфической формы тела камбалой лишь после того, как из икринки вылупляется малек, а также появление изгибов позвоночника, сращение швов в мозговом черепе.
2. Девиации — уклонения, возникающие в процессе морфогенеза органа. Примером может являться развитие сердца в онтогенезе млекопитающих, у которых оно рекапитулирует стадию трубки, двухкамерное и трехкамерное строение, но стадия формирования неполной перегородки, характерной для пресмыкающихся, вытесняется развитием перегородки, построенной и расположенной иначе и характерной только для млекопитающих
3. Архаллаксисы — изменения, обнаруживающиеся на уровне зачатков и выражающиеся в нарушении их расчленения, ранних дифференцировок или в появлении принципиально новых закладок. Классическим примером архаллаксиса является развитие волос у млекопитающих, закладка которых наступает на очень ранних стадиях развития и с самого начала отличается от закладок других придатков кожи позвоночных. В процессе онтогенеза происходит развитие, а часто и замена одних органов другими. Органы зрелого организма называют дефинитивными; органы, развивающиеся и функционирующие только в зародышевом или личиночном развитии, — провизорными. Примерами провизорных органов являются жабры личинок земноводных, первичная почка и зародышевые оболочки высших позвоночных животных (амниот).
Если у двух организмов, находящихся на разных уровнях организации, обнаруживаются органы, которые построены по единому плану, расположены в одинаковом месте и развиваются сходным образом из одинаковых эмбриональных зачатков, то это свидетельствует о родстве данных организмов. Такие органы называют гомологичными. Гомологичные органы часто выполняют одну и ту же функцию (например, сердце рыбы, земноводного, пресмыкающегося и млекопитающего), но в процессе эволюции функции могут и меняться (например, передних конечностей рыб и земноводных, пресмыкающихся и птиц). При обитании неродственных организмов в одинаковых средах у них могут возникать сходные приспособления, которые проявляются в возникновении аналогичных органов. Аналогичные органы выполняют одинаковые функции, строение же их, местоположение и развитие резко различны. Примерами таких органов являются крылья насекомых и птиц, конечности и челюстной аппарат членистоногих и позвоночных.
107. Филогенез нервной системы позвоночных. Филогенетически обусловленные пороки развития головного мозга как результат нарушения онтогенеза.
Нервная система развивается из эктодермы. У всех позвоночных центральная нервная система является производной нервной трубки, передний конец которой становится головным мозгом, а задний — спинным. Образование головного мозга называют кефализацией. Головной мозг всех современных позвоночных животных в эмбриогенезе закладывается вначале из трех мозговых пузырей, и только позже дифференцируется на пять отделов. Головной мозг современных взрослых позвоночных всегда состоит из пяти отделов: переднего, промежуточного, среднего, заднего и продолговатого. Имеются также полости - спинном мозге это спинномозговой канал, а в головном — желудочки мозга. Ткань мозга состоит из серого вещества (скопления нервных клеток) и белого (отростков нервных клеток).
У рыб головной мозг в целом невелик. Слабо развит его передний отдел. Передний мозг не разделен на полушария. Крыша его тонкая, состоит только из эпителиальных клеток и не содержит нервной ткани. Основание переднего мозга включает полосатые тела, от него отходят обонятельные доли. Функционально передний мозг является высшим обонятельным центром. В промежуточном мозге, с которым связаны эпифиз и гипофиз, расположен гипоталамус, являющийся центральным органом эндокринной системы. Средний мозг рыб наиболее развит. Он состоит из двух полушарий и служит высшим зрительным центром. Кроме того, он представляет собой высший интегрирующий отдел головного мозга. Задний мозг содержит мозжечок, осуществляющий регуляцию координации движений. Он развит очень хорошо в связи с перемещением рыб в трехмерном пространстве. Продолговатый мозг обеспечивает связь высших отделов головного мозга со спинным и содержит центры дыхания и кровообращения. Головной мозг такого типа, в котором высшим центром интеграции функций является средний мозг, называют ихтиопсидным.
У земноводных головной мозг также ихтиопсидный. Однако передний мозг их имеет большие размеры и разделен на полушария. Крыша его состоит из нервных клеток, отростки которых располагаются на поверхности. Как и у рыб, больших размеров достигает средний мозг, также представляющий собой высший интегрирующий центр и центр зрения. Мозжечок несколько редуцирован в связи с примитивным характером движений.
Условия наземного существования пресмыкающихся требуют более сложной морфофункциональной организации мозга. Передний мозг — наиболее крупный отдел по сравнению с остальными. В нем особенно развиты полосатые тела. К ним переходят функции высшего интегративного центра. На поверхности крыши впервые появляются островки коры очень примитивного строения, ее называют древней — archicortex. Средний мозг теряет значение ведущего отдела, и относительные размеры его сокращаются. Мозжечок сильно развит благодаря сложности и многообразию движений пресмыкающихся. Головной мозг такого типа, в котором ведущий отдел представлен полосатыми телами переднего мозга, называют зауропсидным.
У млекопитающих — маммалийный тип мозга. Для него характерно сильное развитие переднего мозга за счет коры. В ней располагаются высшие центры зрительного, слухового, осязательного, двигательного анализаторов, а также центры высшей нервной деятельности. Кора имеет очень сложное строение и называется новой корой — neocortex. В ней располагаются не только тела нейронов, но и ассоциативные волокна, соединяющие разные ее участкиПромежуточный мозг, как и у других классов, включает гипоталамус, гипофиз и эпифиз. В среднем мозге располагается четверохолмие в виде четырех бугров. Очень хорошо развит мозжечок.
По мере усиления функций передних отделов головного мозга в филогенезе спинного мозга наблюдается его продольная дифференцировка с образованием утолщений в области отхождения крупных нервов к конечностям и редукция его заднего конца. Так, у рыб спинной мозг равномерно тянется вдоль всего тела. Начиная от земноводных происходит его укорочение сзади. У млекопитающих на заднем конце спинного мозга остается рудимент в виде конечной нити — filum terminale.
Центральная нервная система столь важна для интеграции индивидуального развития человека, что большинство ее врожденных пороков несовместимы с жизнью. Среди пороков спинного мозга, онтогенетические механизмы которых известны, отметим рахисхиз— отсутствие замыкания нервной трубки. Эта аномалия связана с нарушением клеточных перемещений и адгезии в зоне формирования нервной трубки в процессе нейруляции. Аномалия переднего мозга — прозэнцефалия — выражается в нарушении морфогенеза мозга, при котором полушария оказываются неразделенными, а кора — недоразвита. Этот порок формируется на 4-й неделе эмбриогенеза, в момент закладки переднего мозга. Как и предыдущий, он несовместим с жизнью. Часто встречается у мертворожденных при различных хромосомных и генных синдромах. Нарушения дифференцировки коры — агирия (отсутствие извилин) и олигогирия с пахигирией (малое количество утолщенных извилин) — сопровождаются упрощением гистологического строения коры. У детей с такими пороками выявляются грубая олигофрения и нарушение многих рефлексов. Большинство детей умирают в течение первого года жизни.
108. Филогенез нервной системы позвоночных. Филогенетически обусловленные пороки развития головного мозга как результат нарушения онтогенеза.
Нервная система хордовых животных, как и у всех многоклеточных, развивается из эктодермы. Она возникла за счет погружения чувствительных клеток, первоначально лежавших на поверхности тела, под его покровы. Это доказывается и сравнительно-анатомическими, и эмбриологическими данными.
Действительно, у наиболее примитивного представителя хордовых — ланцетника — центральная нервная система, состоящая из нервной трубки, сохранила функции органа чувств: среди клеток, лежащих внутри нее, имеются отдельные светочувствительные образования — глазки Гессе. Кроме того, основные дистантные органы чувств — зрения, обоняния и слуха — образуются у всех позвоночных первоначально как выпячивания передней части нервной трубки.
В эмбриогенезе нервная система формируется вначале всегда в виде полосы утолщенной эктодермы на спинной стороне зародыша, которая впячивается под покровы и замыкается в трубку с полостью внутри — невроцелем. У ланцетника это замыкание еще не полное, поэтому нервная трубка выглядит как желобок. Передний конец ее расширен. Он гомологичен головному мозгу позвоночных. Большинство клеток нервной трубки ланцетника не являются нервными, они выполняют опорные или рецепторные функции.
У всех позвоночных центральная нервная система является производной нервной трубки, передний конец которой становится головным мозгом, а задний — спинным. Образование головного мозга называют кефализацией. Она связана с усилением двигательной активности позвоночных и необходимостью постоянного анализа раздражении, приходящих из внешней среды, в первую очередь с переднего конца тела. Этот процесс сопровождается также дифференциацией органов чувств, особенно дистантных — обоняния, зрения и слуха. Совместная эволюция органов чувств и головного мозга приводит к возникновению динамических координации между обонятельными рецепторами и передним мозгом, зрительными — и средним, слуховыми — и задним. Видимо, поэтому головной мозг всех современных позвоночных животных в эмбриогенезе закладывается вначале из трех мозговых пузырей — переднего, среднего и заднего — и только позже дифференцируется на пять отделов. Вероятно, предки позвоночных имели более простой головной мозг, развивающийся на основе трех мозговых пузырей.
Головной мозг современных взрослых позвоночных всегда состоит из пяти отделов: переднего, промежуточного, среднего, заднего и продолговатого. Внутри головного и спинного мозга расположена общая полость, соответствующая невроцелю. В спинном мозге это спинномозговой канал, а в головном — желудочки мозга. Ткань мозга состоит из серого вещества (скопления нервных клеток) и белого (отростков нервных клеток).
Во всех отделах головного мозга различают мантию, располагающуюся над желудочками, и основание, лежащее под ними. В прогрессивной эволюции головного мозга проявляется постепенное усиление роли его передних отделов и мантии по сравнению с задними и основанием.
У рыб головной мозг в целом невелик. Слабо развит его передний отдел. Передний мозг не разделен на полушария. Крыша его тонкая, состоит только из эпителиальных клеток и не содержит нервной ткани. Основание переднего мозга включает полосатые тела, от него отходят обонятельные доли. Функционально передний мозг является высшим обонятельным центром.
В промежуточном мозге, с которым связаны эпифиз и гипофиз, расположен гипоталамус, являющийся центральным органом эндокринной системы. Средний мозг рыб наиболее развит. Он состоит из двух полушарий и служит высшим зрительным центром. Кроме того, он представляет собой высший интегрирующий отдел головного мозга. Задний мозг содержит мозжечок, осуществляющий регуляцию координации движений. Он развит очень хорошо в связи с перемещением рыб в трехмерном пространстве. Продолговатый мозг обеспечивает связь высших отделов головного мозга со спинным и содержит центры дыхания и кровообращения. Головной мозг такого типа, в котором высшим центром интеграции функций является средний мозг, называют ихтиопсидным.
У земноводных головной мозг также ихтиопсидный. Однако передний мозг их имеет большие размеры и разделен на полушария. Крыша его состоит из нервных клеток, отростки которых располагаются на поверхности. Как и у рыб, больших размеров достигает средний мозг, также представляющий собой высший интегрирующий центр и центр зрения. Мозжечок несколько редуцирован в связи с примитивным характером движений.
Условия наземного существования пресмыкающихся требуют более сложной морфофункциональной организации мозга. Передний мозг — наиболее крупный отдел по сравнению с остальными. В нем особенно развиты полосатые тела. К ним переходят функции высшего интегративного центра. На поверхности крыши впервые появляются островки коры очень примитивного строения, ее называют древней — archicortex. Средний мозг теряет значение ведущего отдела, и относительные размеры его сокращаются. Мозжечок сильно развит благодаря сложности и многообразию движений пресмыкающихся. Головной мозг такого типа, в котором ведущий отдел представлен полосатыми телами переднего мозга, называют зауропсидным.
У млекопитающих — маммалийный тип мозга. Для него характерно сильное развитие переднего мозга за счет коры, которая развивается на основе небольшого островка коры пресмыкающихся и становится интегрирующим центром мозга. В ней располагаются высшие центры зрительного, слухового, осязательного, двигательного анализаторов, а также центры высшей нервной деятельности. Кора имеет очень сложное строение и называется новой корой — neocortex. В ней располагаются не только тела нейронов, но и ассоциативные волокна, соединяющие разные ее участки. Характерным является также наличие комиссуры между обоими полушариями, в которой располагаются волокна, связывающие их воедино. Промежуточный мозг, как и у других классов, включает гипоталамус, гипофиз и эпифиз. В среднем мозге располагается четверохолмие в виде четырех бугров. Два передних связаны со зрительным анализатором, два задних—со слуховым. Очень хорошо развит мозжечок.
По мере усиления функций передних отделов головного мозга в филогенезе спинного мозга наблюдается его продольная дифференцировка с образованием утолщений в области отхождения крупных нервов к конечностям и редукция его заднего конца.
Так, у рыб спинной мозг равномерно тянется вдоль всего тела. Начиная от земноводных происходит его укорочение сзади. У млекопитающих на заднем конце спинного мозга остается рудимент в виде конечной нити — filum terminale. Нервы, идущие к заднему концу тела, проходят по позвоночному каналу самостоятельно, образуя так называемый конский хвост — cauda equina.
Основные этапы эволюции центральной нервной системы отражаются и в онтогенезе человека. На стадии нейруляции закладывается нервная пластинка, превращающаяся в желобок и затем в трубку. Передний конец трубки образует сначала три мозговых пузыря: передний (I), средний (II) и задний (III). Вслед за этим передний пузырь подразделяется на два, дифференцирующихся на передний (7) и промежуточный (2) мозг — telencephalon, diencephalon. Средний мозговой пузырь развивается в средний (3) мозг — mesencephalon, a задний — в задний (4) мозг — metencephalon — и продолговатый (5) мозг — medulla oblongata.
Задний конец спинного мозга редуцируется, превращаясь в терминальную нить. Позже скорости роста спинного мозга и позвоночника оказываются разными, и к моменту рождения конец спинного мозга оказывается на уровне третьего, а у взрослого человека — уже на уровне первого поясничного позвонка.
Головной мозг рано начинает развиваться по пути, характерному для млекопитающих и человека. Первично почти прямая нервная трубка резко изгибается в области будущего продолговатого и среднего мозга. На этом фоне большие полушария переднего мозга растут с особенно большой скоростью. В результате головной мозг оказывается расположенным над лицевым черепом. Дифференцировка коры приводит к развитию извилин, борозд и формированию высших сенсорных и двигательных центров, в том числе центров письменной и устной речи и др., характерных только для человека.
Центральная нервная система столь важна для интеграции индивидуального развития человека, что большинство ее врожденных пороков несовместимы с жизнью. Среди пороков спинного мозга, онтогенетические механизмы которых известны, отметим рахисхиз,нли платиневрию,— отсутствие замыкания нервной трубки. Эта аномалия связана с нарушением клеточных перемещений и адгезии в зоне формирования нервной трубки в процессе нейруляции. Аномалия переднего мозга — прозэнцефалия — выражается в нарушении морфогенеза мозга, при котором полушария оказываются неразделенными, а кора — недоразвита. Этот порок формируется на 4-й неделе эмбриогенеза, в момент закладки переднего мозга. Как и предыдущий, он несовместим с жизнью. Часто встречается у мертворожденных при различных хромосомных и генных синдромах.
Нарушения дифференцировки коры — агирия (отсутствие извилин) и олигогирия с пахигирией (малое количество утолщенных извилин) — сопровождаются упрощением гистологического строения коры. У детей с такими пороками выявляются грубая олигофрения и нарушение многих рефлексов. Большинство детей умирают в течение первого года жизни.
109. Филогенез кровеносной системы позвоночных. Филогенетически обусловленные пороки развития сердца и сосудов как результат нарушения онтогенеза.
Необходимым условием существования высокоорганизованных крупных многоклеточных организмов является наличие жидкой подвижной внутренней среды, которая обеспечивает интеграцию организма в целостную систему, выполняя транспортные функции. Эти функции являются основными для кровеносной системы. Конкретная функция кровеносной системы зависит от того, что она транспортирует: питательные вещества, кислород, углекислый газ, другие продукты диссимиляции или гормоны. Кровеносная система всех хордовых замкнутая и состоит из двух основных артериальных сосудов: брюшной и спинной аорт. По брюшной аорте венозная кровь продвигается кпереди, обогащается кислородом в органах дыхания, а по спинной — кзади. Из спинной аорты кровь через систему капилляров возвращается по венам в брюшную аорту. Брюшная аорта или ее часть, периодически сокращаясь, проталкивает кровь по сосудам.
Эволюция общего плана строения кровеносной системы хордовых
У ланцетника кровеносная система наиболее проста. Круг кровообращения один. По брюшной аорте венозная кровь поступает в приносящие жаберные артерии, которые по количеству соответствуют числу межжаберных перегородок (до 150 пар), где и обогащается кислородом.
По выносящим жаберным артериям кровь поступает в корни спинной аорты, расположенные симметрично с двух сторон тела. Они продолжаются как вперед, неся артериальную кровь к головному мозгу, так и назад. Передние ветви этих двух сосудов являются сонными артериями. На уровне заднего конца глотки задние ветви образуют спинную аорту, которая разветвляется на многочисленные артерии, направляющиеся к органам и распадающиеся на капилляры.
После тканевого газообмена кровь поступает в парные передние или задние кардинальные вены, расположенные симметрично. Передняя и задняя кардинальные вены с каждой стороны впадают в кювьеров проток. Оба кювьеровых протока впадают с двух сторон в брюшную аорту. От стенок пищеварительной системы венозная кровь оттекает по воротной вене печени в печеночный вырост, где формируется система капилляров. Затем капилляры вновь собираются в венозный сосуд — печеночную вену, по которой кровь поступает в брюшную аорту.
Таким образом, несмотря на простоту кровеносной системы в целом, уже у ланцетника имеются основные магистральные артерии, характерные для позвоночных, в том числе для человека: это брюшная аорта, преобразующаяся позже в сердце, восходящую часть дуги аорты и корень легочной артерии; спинная аорта, становящаяся позже собственно аортой, и сонные артерии. Основные вены, имеющиеся у ланцетника, также сохраняются у более высокоорганизованных животных. Так, передние кардинальные вены станут позже яремными венами, правый кювьеров проток преобразуется в верхнюю полую вену, а левый, сильно редуцировавшись, — в коронарный синус сердца. Для того чтобы понять, как это происходит, необходимо сопоставить кровеносные системы всех классов позвоночных животных.
Более активный образ жизни рыб предполагает более интенсивный метаболизм. В связи с этим на фоне олигомеризации их артериальных жаберных дуг в конечном счете до четырех пар в них отмечается высокая степень дифференцировки: жаберные сосуды распадаются на капилляры, пронизывающие жаберные лепестки. В процессе интенсификации сократительной функции брюшной аорты часть ее преобразовалась в двухкамерное сердце, состоящее из предсердия и желудочка и располагающееся под нижней челюстью, рядом с жаберным аппаратом. В остальном кровеносная система рыб соответствует строению ее у ланцетника.
В связи с выходом земноводных на сушу и появлением легочного дыхания у них возникает два круга кровообращения. Соответственно этому в строении сердца и артерий появляются приспособления, направленные на разделение артериальной и венозной крови. Перемещение земноводных в основном за счет парных конечностей, а не хвоста обусловливает изменения в венозной системе задней части туловища.
Сердце амфибий расположено каудальнее, чем у рыб, рядом с легкими; оно трехкамерное, но, как и у рыб, от правой половины единственного желудочка начинается единственный сосуд — артериальный конус, разветвляющийся последовательно на три пары сосудов:
кожно-легочные артерии, дуги аорты и сонные артерии. Как и у всех более высокоорганизованных классов, в правое предсердие впадают вены большого круга, несущие венозную кровь, в левое— малого с артериальной кровью. При сокращении предсердий в желудочек, внутренняя стенка которого снабжена большим количеством мышечных перекладин, одновременно попадают обе порции крови. Полного их смешения из-за своеобразного строения стенки желудочка не происходит, поэтому при его сокращении первая порция венозной крови поступает в артериальный конус и с помощью спирального клапана, находящегося там, направляется в кожно-легочные артерии. Кровь из середины желудочка, смешанная, поступает таким же образом в дуги аорты, а оставшееся небольшое количество артериальной крови, последней попадающей в артериальный конус, направляется в сонные артерии.
Две дуги аорты, несущие смешанную кровь, огибают сердце и пищевод сзади, образуя спинную аорту, снабжающую все тело, кроме головы, смешанной кровью. Задние кардинальные вены сильно редуцируются и собирают кровь только с боковых поверхностей туловища. Функционально их замещает возникшая заново задняя полая вена, собирающая кровь в основном из задних конечностей. Она располагается рядом со спинной аортой и, находясь позади печени, вбирает в себя печеночную вену, которая у рыб впадала непосредственно в венозный синус сердца. Передние кардинальные вены, обеспечивая отток крови от головы, называют теперь яремными венами, а кювьеровы потоки, в которые они впадают вместе с подключичными венами, — передними полыми венами.
В кровеносной системе пресмыкающихся возникают следующие прогрессивные изменения: в желудочке их сердца имеется неполная перегородка, затрудняющая смешение крови, поступающей из правого и левого предсердий; от сердца отходит не один, а три сосуда, образовавшихся в результате разделения артериального ствола. Из левой половины желудочка начинается правая дуга аорты, несущая артериальную кровь, а из правой — легочная артерия с венозной кровью. Из середины желудочка, в области неполной перегородки, начинается левая дуга аорты со смешанной кровью. Обе дуги аорты, как и у предков, срастаются позади сердца, трахеи и пищевода в спинную аорту, кровь в которой смешанная, но более богата кислородом, чем у земноводных, в связи с тем что до слияния сосудов только по левой дуге течет смешанная кровь. Кроме того, сонные и подключичные артерии с обеих сторон берут начало от правой дуги аорты, в результате чего артериальной кровью снабжается не только голова, но и передние конечности. В связи с появлением шеи сердце располагается еще более каудально, чем у земноводных. Венозная система пресмыкающихся принципиально не отличается от системы вен земноводных.
Прогрессивные изменения кровеносной системы млекопитающих сводятся к полному разделению венозного и артериального кровотоков. Это достигается, во-первых, завершенной четырехкамерностью сердца и, во-вторых, редукцией правой дуги аорты и сохранением только левой, начинающейся от левого желудочка. В результате все органы млекопитающих снабжаются артериальной кровью. В венах большого круга кровообращения также обнаруживаются прогрессивные изменения: возникла безымянная вена, объединяющая левые яремную и подключичную вены с правыми, в результате чего остается лишь одна передняя полая вена, располагающаяся справа. Левый кювьеров проток в виде рудиментарного сосуда sinus coronarius теперь собирает венозную кровь только от миокарда, а непарная и полунепарная вены — рудименты задних кардинальных вен, имеют существенное значение в основном в случаях формирования обходных путей венозного оттока через кава-кавальные анастомозы, формируемые ими.
В эмбриональном развитии млекопитающих и человека рекапитулируют закладки сердца и основных кровеносных сосудов предковых классов.
Сердце закладывается на первых этапах развития в виде недифференцированной брюшной аорты, которая за счет изгибания, появления в просвете перегородок и клапанов, становится последовательно двух-, трех- и четырехкамерным. Однако рекапитуляции здесь неполны в связи с тем, что межжелудочковая перегородка млекопитающих формируется иначе и из другого материала по сравнению с рептилиями. Поэтому можно считать, что четырехкамерное сердце млекопитающих формируется на базе трехкамерного сердца, а межжелудочковая перегородка является новообразованием, а не результатом доразвития перегородки пресмыкающихся. Таким образом, в филогенезе сердца позвоночных проявляется девиация: в процессе морфогенеза этого органа у млекопитающих рекапитулируют ранние филогенетические стадии, а затем развитие его идет в ином направлении, характерном лишь для этого класса.
Интересно, что место закладки и положение сердца в филогенетическом ряду позвоночных полностью рекапитулируют у млекопитающих и человека. Так, закладка сердца у человека осуществляется на 20-е сутки эмбриогенеза, как у всех позвоночных, позади головы. Позже за счет изменения пропорций тела, появления шейной области, смещения легких в грудную полость осуществляется и перемещение сердца в переднее средостение.
Нарушения развития сердца могут выражаться как в возникновении аномалий строения, так и места его положения. Возможно сохранение к моменту рождения двухкамерного сердца. Этот порок совершенно не совместим с жизнью.
Чаще встречаются дефекты межпредсердной перегородки (1 случай на 1000 рождений), межжелудочковой перегородки (2,5—5 случаев на 1000 рождений), вплоть до трехкамерного сердца с одним общим желудочком. Известен и такой порок, как шейная эктопия сердца, при которой оно находится в шейной области. Этот порок связывают с задержкой сердца в области его первоначальной закладки. При этом ребенок обычно погибает сразу после рождения. Перечисленные пороки сердца наиболее часто встречаются не в изолированном виде, а в комплексе с другими аномалиями сердца, сосудов, а нередко и других органов. Это свидетельствует о том, что в морфогенезе сердца большое значение имеют онтогенетические корреляции. Состояние больных при таких пороках зависит от того, насколько сильно нарушается гемодинамика и осуществляется смешение крови в кровеносном русле.
Филогенез артериальных жаберных дуг
В связи с тем что основные артериальные сосуды у млекопитающих и человека формируются на базе закладок жаберных артерий, проследим их эволюцию в филогенетическом ряду позвоночных. В эмбриогенезе абсолютного большинства позвоночных закладывается шесть пар артериальных жаберных дуг, соответствующих шести парам висцеральных дуг черепа. В связи с тем что две первые пары висцеральных дуг включаются в состав лицевого черепа, две первые артериальные жаберные дуги быстро редуцируются. Оставшиеся четыре пары функционируют у рыб как жаберные артерии. У наземных позвоночных 3-я пара жаберных артерий теряет связь с корнями спинной аорты и несет кровь к голове, становясь сонными артериями. Сосуды 4-й пары достигают наибольшего развития и вместе с участком корня спинной аорты во взрослом состоянии становятся дугами аорты — основными сосудами большого круга кровообращения.
У земноводных и пресмыкающихся оба сосуда развиты и принимают участие в кровообращении. У млекопитающих также закладываются оба сосуда 4-й пары, а позже правая дуга аорты редуцируется таким образом, что от нее остается лишь небольшой рудимент— плечеголовной ствол. Пятая пара артериальных дуг в связи с тем, что она функционально дублирует четвертую, редуцируется у всех наземных позвоночных, кроме хвостатых амфибий. Шестая пара, которая снабжает венозной кровью кроме жабр еще и плавательный пузырь, у кистеперых рыб становится легочной артерией.
В эмбриогенезе человека рекапитуляции артериальных жаберных дуг происходят с особенностями: все шесть пар дуг никогда не существуют одновременно. В то время, когда две первые дуги закладываются, а затем перестраиваются, последние пары сосудов еще не начинают формироваться. Кроме того, пятая артериальная дуга уже закладывается в виде рудиментарного сосуда, присоединенного обычно к 4-й паре, и редуцируется очень быстро.
Из атавистических пороков развития сосудов, развивающихся из артериальных жаберных дуг, остановимся на следующих: с частотой 1 случай на 200 вскрытии детей, умерших от врожденных пороков сердца, встречается персистирование обеих дуг аорты 4-й пары. При этом обе дуги, так же как у земноводных или пресмыкающихся, срастаются позади пищевода и трахеи, образуя нисходящую часть спинной аорты. Порок проявляется нарушением глотания и удушьем. Несколько чаще (2,8 случая на 200 вскрытии) встречается нарушение редукции правой дуги аорты с редукцией левой. Эта аномалия часто клинически не проявляется.
Наиболее частый порок (0,5—1,2 случая на 1000 новорожденных) — персистирование артериального, или боталлова, протока представляющего собой часть корня спинной аорты между 4-й и'6-и парами артерий слева. Проявляется сбросом артериальной крови из большого круга кровообращения в малый. Очень тяжелый порок развития — персистирование первичного эмбрионального ствола, в результате которого из сердца выходит только один сосуд, располагающийся обычно над дефектом в межжелудочковой перегородке. Он обычно заканчивается смертью ребенка. Нарушение дифференцировки первичного эмбрионального ствола может привести к такому пороку развития, как транспозиция сосудов — отхождение аорты от правого желудочка, а легочного ствола — от левого, что встречается в 1 случае на 2500 новорожденных. Этот порок обычно несовместим с жизнью.
Рекапитуляции проявляются и в эмбриональном развитии крупных вен человека. При этом возможно формирование атавистических пороков развития. Среди пороков развития венозного русла укажем на возможность персистирования двух верхних полых вен. Если обе они впадают в правое предсердие, аномалия клинически не проявляется. При впадении левой полой вены в левое предсердие происходит сброс венозной крови в большой круг кровообращения. Иногда обе полые вены впадают в левое предсердие. Такой порок несовместим с жизнью. Данные аномалии встречаются с частотой 1% от всех врожденных пороков сердечно-сосудистой системы.
Очень редкая врожденная аномалия — неразвитие нижней полой вены. Отток крови от нижней части туловища и ног осуществляется в этом случае через коллатерали непарной и полунепарной вен, являющихся рудиментами задних кардинальных вен.
110. Филогенез мочеполовой системы позвоночных. Филогенетически обусловленные пороки развития мочеполовой системы как результат нарушения онтогенеза.
Выделительная и половая системы в целом выполняют разные функции. Однако их рассматривают в едином комплексе в связи с единством эмбрионального развития и первичной функциональной связью со вторичной полостью тела — целбмом. В эмбриогенезе закладка мочеполовой системы — нефрогонотом — формируется в области ножки сомита в непосредственном контакте с целомом. Половые железы у всех хордовых животных располагаются в целоме. Продукты диссимиляции у всех целоми-ческих животных, в том числе и низших хордовых, также поступают в целом. Наиболее простой путь выведения во внешнюю среду как половых клеток, так и продуктов диссимиляции — через общий канал, начинающийся воронкой в целоме и заканчивающийся выделительной порой на покровах. Таковы взаимоотношения половых желез и выделительных каналов у многих беспозвоночных и, вероятно, у древних предков хордовых.
Эволюция почки
Органами выделения позвоночных являются почки — парные компактные органы, структурная единица которых представлена нефроном. В наиболее примитивном виде это — воронка, открывающаяся в целом и соединенная с выделительным канальцем, который впадает в общий выводной проток — мочеточник. В филогенезе позвоночных почка прошла три этапа эволюции: предпочка — головная, или пронефрос; первичная почка—туловищная, или мезонефрос, и вторичная почка - тазовая, или метанефрос.
Предпочка полностью развивается и функционирует как самостоятельный орган у личинок рыб и земноводных. Она находится на переднем конце тела, состоит из 2—12 нефронов, воронки которых открыты в целом, а выводные канальцы впадают в пронефрический канал, который соединен с клоакой. Предпочка имеет сегментарное строение. Продукты диссимиляции фильтруются в целом из кровеносных сосудов, которые поблизости от нефронов формируют клубочки.
У взрослых рыб и земноводных кзади от предпочек, в туловищных сегментах тела, формируются первичные почки, содержащие до нескольких сотен нефронов. В ходе онтогенеза нефроны увеличиваются в количестве за счет их почкования друг от друга с последующей дифференцировкой. Они вступают в связь с кровеносной системой, формируя капсулы почечных клубочков. Капсулы имеют вид двустенных чаш, в которых располагаются сосудистые клубочки, благодаря чему продукты диссимиляции могут поступать из крови непосредственно в нефрон. Некоторые нефроны первичной почки сохраняют связь с целомом через воронки, другие — утрачивают её.
Выделительные канальцы удлиняются и в них осуществляется обратное всасывание в кровь воды, глюкозы и других веществ, в связи с чем концентрация продуктов диссимиляции в моче повышается. Однако воды с мочой теряется много, поэтому животные, обладающие такой почкой, могут обитать только в водной или влажной среде. Первичная почка сохраняет признаки метамерного строения.
У пресмыкающихся и млекопитающих возникают вторичные почки. Они закладываются в тазовом отделе тела и содержат сотни тысяч нефронов наиболее совершенного строения. У новорожденного ребенка в почке их насчитывается около 1 млн. Они образуются за счет многократного ветвления развивающихся нефронов. Нефроны не имеют воронки и, таким образом, теряют полностью связь с целомом. Канадец нефрона удлиняется, теснее контактирует с кровеносной системой, а у млекопитающих дифференцируется на проксимальный и дистальный участки, между которыми появляется еще и так называемая петля Генле.
Такое строение нефрона обеспечивает не только полноценную фильтрацию плазмы крови в капсуле, но и, что более важно, эффективное обратное всасывание в кровь воды, глюкозы, гормонов, солей и других необходимых организму веществ. В результате концентрация продуктов диссимиляции в моче, выделяемой вторичными почками, велика, а само ее количество — мало. У человека, например, за сутки в капсулах нефронов обеих почек фильтруется около 150 л плазмы крови, а мочи выделяется около 2 л. Это позволяет животным, обладающим вторичными почками, быть более независимыми от водной среды и заселять засушливые участки земли. У пресмыкающихся вторичные почки на протяжении всей жизни сохраняются на месте их первоначальной закладки — в тазовой области. В них прослеживаются черты первичного метамерного строения.
Почки млекопитающих располагаются в поясничной области, и у большинства из них внешняя сегментация не выражена. В онтогенезе человека обнаруживается выраженная рекапитуляция в развитии почки: закладывание вначале про-, затем мезо-, а позже метанефроса. Последний развивается в тазовой области, а затем за счет различий в скоростях роста позвоночника, таза и органов брюшной полости перемещается в поясничную область. У пятинедельного зародыша можно обнаружить сосуществование предпочки, первичной, а также зачатков вторичной почки.
На начальных этапах развития почка человека сегментирована. Позже ее поверхность сглаживается и метамерность сохраняется лишь во внутреннем строении в виде почечных пирамид. Пороки развития почек у человека, основанные на их филогенезе, многообразны. Сохранение мезонефроса и одностороннее отсутствие вторичной почки описаны пока только у мышей, хотя в принципе такая аномалия возможна и у человека. Относительно часто встречается сегментированная вторичная почка, имеющая один или даже несколько мочеточников; возможно и полное ее удвоение. Часто наблюдается тазовое расположение почки, связанное с нарушением ее перемещения на 2—4-м месяцах зародышевого развития.
Эволюция половых желез
Половые железы у всех позвоночных развиваются в виде парных складок части нефрогонотома в области ножки сомита. Половые складки вдаются в полость тела и оказываются подвешенными на брыжейке. Первичные половые клетки обособляются у зародышей очень рано — уже на стадии гаструляции. Вначале они обнаруживаются в составе презумптивной эктодермы головного конца эмбриона, затем попадают в энтодерму, откуда активно перемещаются в половые складки. Здесь дифференцирующийся эпителий половой железы, включающий в себя первичные половые клетки, объединяется с соединительнотканной стромой в виде шнуров. Такая гонада индифферентна в половом отношении и может развиваться в дальнейшем как в семенник, так и в яичник в зависимости от генетических и эпигенетических факторов дифференцировки пола.
У хрящевых рыб строение половых желез самок и самцов в значительной степени сходно. У всех остальных позвоночных яичник всегда имеет фолликулярное строение, т.е. содержит пузырьки — фолликулы, в каждом из которых находится одна будущая яйцеклетка. При созревании яйцеклетки стенка фолликула разрывается, и она поступает вначале в брюшную полость, а затем в яйцевод. Семенники содержат семенные трубочки, которые соединены с семявыносящими каналами, по которым зрелые сперматозоиды выходят во внешнюю среду.
Индифферентность развивающейся половой железы позвоночных называют первичным гермафродитизмом. Он эволюционно связан, вероятно, с гермафродитизмом древних предков позвоночных. Доказательством этого может служить наличие у наиболее примитивных современных позвоночных — круглоротых — таких половых желез, передняя часть которых является яичником, а задняя — семенником. У самок некоторых земноводных на протяжении всей жизни сохраняется рудиментарный участок индифферентной половой железы. Зародыш человека до определенного возраста имеет недифференцированные половые железы, которые в зависимости от различных факторов становятся либо семенниками, либо яичниками. Нарушение их дифференцировки может привести к возникновению ovotestis, в котором сочетаются элементы семенника и яичника. У детей, имеющих ovotestis, обнаруживаются признаки гермафродитизма и в наружных половых органах.
У всех позвоночных с непостоянной температурой тела половые железы находятся в брюшной полости. У большинства млекопитающих мужские гонады перемещаются через паховой канал в мошонку, где температура всегда несколько ниже. Вероятно, пониженная температура более благоприятна для сперматогенеза, обеспечивая более низкий уровень спонтанных мутаций в созревающих сперматозоидах.
У человека семенники, закладываясь в брюшной полости, перемещаются через паховой канал и к 8-му месяцу внутриутробного развития оказываются в мошонке. У 2,2 % мальчиков пубертатного возраста обнаруживаются различные формы крипторхизма — неопущения яичек, которые при этом обычно недоразвиты, а часть семявыносящих канальцев заменена соединительной тканью. Для предотвращения бесплодия таким мальчикам необходимо хирургическое низведение яичек в мошонку в раннем возрасте.
Эволюция мочеполовых протоков
В эмбриогенезе всех позвоночных при развитии предпочки вдоль тела, от головного конца к клоаке, закладывается канал, по которому продукты диссимиляции из нефронов поступают во внешнюю среду. Это пронефрический канал. При развитии первичной почки этот канал либо расщепляется на два канала, идущих параллельно, либо второй канал образуется в продольном утолщении стенки первого. Один из них — вольфов — вступает в связь с нефронами первичной почки. Другой — мюллеров — срастается передним концом с одним из нефронов предпочки и образует яйцевод, открывающийся передним концом в целом широкой воронкой, а задним — впадающий в клоаку.
Вне зависимости от пола у всех позвоночных обязательно формируются как вольфов, так и мюллеров каналы, однако судьба их различна как у разных полов, так и у представителей разных классов. У самок рыб и земноводных вольфов канал всегда выполняет функцию мочеточника, а мюллеров — яйцевода. У самцов мюллеров канал редуцируется и обе функции — половую и выделительную — выполняет вольфов канал. Семенные канальцы при этом впадают в почку, а сперматозоиды при оплодотворении поступают в воду вместе с мочой.
У пресмыкающихся и млекопитающих большая часть вольфова канала не принимает участия в выведении мочи и только его наиболее каудальная часть в области впадения в клоаку образует выпячивание, становящееся мочеточником вторичной почки. Сам же вольфов канал у самцов выполняет функцию семяизвергательного канала. Мюллеров канал у них подвергается редукции. У самок вольфов канал редуцируется (за исключением его каудальной части, формирующей мочеточник), а мюллеров — становится яйцеводом. У плацентарных млекопитающих мюллеров канал дифференцируется на собственно яйцевод, матку и влагалище. Будучи парным образованием, как и все элементы половой системы, мюллеров канал сохраняет парность строения у яйцекладущих и частично у сумчатых млекопитающих, у которых имеется два влагалища, две матки и два яйцевода. В дальнейшей эволюции происходит срастание мюллеровых каналов с образованием одного влагалища и матки, которая может быть либо двойной, как у многих грызунов, либо двураздельной, как у хищных; либо двурогой, как у насекомоядных и китообразных, либо простой, как у приматов и человека.
Соответственно дифференцировкам мюллерова канала самок у самцов пресмыкающихся и млекопитающих развиваются копулятивные органы. У большинства пресмыкающихся, а также у сумчатых млекопитающих они парные. У плацентарных с одним влагалищем копулятивный орган непарный, но в его развитии обнаруживается срастание парных зачатков.
В эмбриогенезе человека закладываются парные вольфовы и мюллеровы каналы. Позже в зависимости от пола происходит их редукция. Рудимент мюллерова канала у мужчин располагается в предстательной железе и называется мужской маточкой — utriculus masculinus. Канальцы передней части первичной почки у них вступают в связь с семенниками и преобразуются в придаток семенника — эпидидимис. У плодов женского пола возможно нарушение редукции вольфовых каналов, которые располагаются по бокам от влагалища. Эта аномалия опасна возможностью образования кист и злокачественного перерождения. Распространенными пороками развития являются также различные формы удвоения матки (1 случай на 1000 перинатальных вскрытии). Они развиваются как результат нарушения срастания мюллеровых каналов. Нарушение срастания парных зачатков полового члена в эмбриогенезе человека может привести к формированию такого порока развития, как его удвоение.
111. Популяционная структура человечества. Демы, изоляты, неизолированные популяции. Распределение и частота наследственных заболеваний в разных популяциях людей.
ПОПУЛЯЦИЯ ЛЮДЕЙ. ДЕМ, ИЗОЛЯТ
Размножение человека осуществляется половым путем, а репродуктивные ареалы в той или иной степени ограничены определенной группой населения. Это позволяет выделить в человечестве сообщества, аналогичные популяциям в биологическом понимании этого термина. В антропогенетике популяцией называют группу людей, занимающих общую территорию и свободно вступающих в брак. Изоляционные барьеры, препятствующие заключению брачных союзов, нередко носят выраженный социальный характер (например, различия в вероисповедании). Благодаря этому в формировании популяций людей главную роль играет не общность территории, а социальные факторы.
Размер, уровень рождаемости и смертности, возрастной состав, экономическое состояние, уклад жизни являются демографическими показателями популяций людей. Генетически они характеризуются генофондами (аллелофондами). Демографические показатели оказывают серьезное воздействие на состояние генофондов человеческих популяций, главным образом через структуру браков. Большое значение в определении структуры браков имеет размер группы.
Популяции из 1500—4000 человек называют демами, популяции численностью до 1500 человек — изолятами. Для демов и изолятов типичен относительно низкий естественный прирост населения — соответственно порядка 20% и не более 25% за поколение. Частота внутригрупповых браков в них составляет 80—90% и свыше 90%, а приток лиц из других групп сохраняется на уровне 1—2% и менее 1%. В силу высокой частоты внутригрупповых браков члены изолятов, просуществовавших четыре поколения (примерно 100 лет) и более, являются не менее чем троюродными братьями и сестрами (сибсами).
В больших по размерам популяциях распределение аллелей отдельных генов в генотипах индивидуумов последовательных поколений подчиняется закону Харди — Вайнберга. Это используют в медико-генетической практике для расчета доли гетерозигот — носителей определенного рецессивного аллеля. Так, в Швеции в 1965—1974 гг. страдающие фенилкетонурией встречались с частотой примерно 1 : 40000. Исходя из закона Харди — Вайнберга, по локусу, представленному двумя аллелями, три возможных генотипа (AlA1, A1A2 и А2А2) распределяются с частотой р2, 2pq, q2. Следовательно, q2 = 1/40000, a q = 1/200. Частота доминантного аллеля нормального обмена фенилаланина р=1—q=l— 1/200 = 199/200. Тогда частота гетерозигот 2pq = 2 х (1/200) • (199/200) = 2 • (199/40000). При найденных частотах доминантного и рецессивного аллелей популяция численностью 40 000 человек содержит одного больного фенилкетонурией (А2А2) и 400 носителей неблагоприятного аллеля в гетерозиготном состоянии (A1A2). Остальные члены популяции гомозиготны по благоприятному доминантному аллелю (AlA1). Ниже приведены данные о частоте гетерозиготного носительства и соответствующей ей частоте рецессивных гомозигот с фенотипическим проявлением определенного аллеля.
Встречаемость гомозигот (в пересчете на число членов популяции) |
Встречаемость гетерозигот (в пересчете на число членов популяции) |
1 : 10 |
1 : 2,3 |
1 : 100 |
1 : 5.6 |
1 : 1000 |
1 : 16 |
1 : 10 000 |
1 : 51 |
1 : 100 000 |
1 : 159 |
1 : 1 000 000 |
1 : 501 |
Даже по редким рецессивным аллелям количество гетерозигот оказывается достаточно высоким, чтобы это учитывалось при медико-генетическом консультировании вступающих в брак.
Генетическое разнообразие в популяциях людей
Человечеству свойствен высокий уровень наследственного разнообразия, что проявляется в многообразии фенотипов. Люди отличаются друг от друга цветом кожных покровов, глаз, волос, формой носа и ушной раковины, рисунком эпидермальных гребней на подушечках пальцев и другими сложными признаками. Выявлены многочисленные варианты отдельных белков, различающиеся по одному или нескольким аминокислотным остаткам и, следовательно, функционально. Белки являются простыми признаками и прямо отражают генетическую конституцию организма. У людей не совпадают группы крови по системам эритроцитарных антигенов «резус», АВ0, MN. Известно более 130 вариантов гемоглобина, более 70 вариантов фермента глюкозо-6-фосфатдегидрогеназы (Г6ФД), который участвует в бескислородном расщеплении глюкозы в эритроцитах. В целом не менее 30% генов, контролирующих у человека синтез ферментных и других белков, имеют несколько аллельных форм. Частота встречаемости разных аллелей одного гена варьирует.
Так, из многих вариантов гемоглобина лишь четыре обнаруживаются в некоторых популяциях в высокой концентрации: HbS (тропическая Африка, Средиземноморье), НЬС (Западная Африка), HbD (Индия), НЬЕ (Юго-Восточная Азия). Концентрация других аллелей гемоглобина повсеместно не превышает, видимо, 0,01—0,0001. Вариабельность распространенности аллелей в популяциях людей зависит от действия элементарных эволюционных факторов. Важная роль принадлежит мутационному процессу, естественному отбору, генетико-автоматическим процессам, миграциям.
Мутационный процесс создает новые аллели. И в человеческих популяциях он действует ненаправленно, случайным образом. В силу этого отбор не приводит к выраженному преобладанию концентрации одних аллелей над другими. В достаточно большой популяции, где каждая пара родителей из поколения в поколение дает двух потомков вероятность сохранения новой нейтральной мутации через 15 поколений составляет всего 1/9.
Все многообразие вариантов белков, отражающее разнообразие аллелей в генофонде человечества, можно разделить на две группы. К одной из них относятся редкие варианты, встречающиеся повсеместно с частотой менее 1%. Появление их объясняется исключительно мутационным процессом. Вторую группу составляют варианты, обнаруживаемые относительно часто в избранных популяциях. Так, в примере с гемоглобинами к первой группе относятся все варианты, кроме HbS, HbC, HbD и HbE. Длительные различия в концентрации отдельных аллелей между популяциями, сохранение в достаточно высокой концентрации нескольких аллелей в одной популяции зависят от действия естественного отбора или дрейфа генов.
К межпопуляционным различиям в концентрации определенных аллелей приводит стабилизирующая форма естественного отбора. Неслучайное распределение по планете аллелей эритроцитарных антигенов АВ0 может быть, например, обусловлено различной выживаемостью лиц, отличающихся по группе крови, в условиях частых эпидемий особо опасных инфекций. Области сравнительно низких частот аллеля I0 и относительно высоких частот аллеля IB в Азии примерно совпадают с очагами чумы. Возбудитель этой инфекции имеет Н-подобный антиген. Это делает людей с группой крови О особенно восприимчивыми к чуме, так как они, имея антиген Н, не способны вырабатывать противочумные антитела в достаточном количестве. Указанному объяснению соответствует факт, что относительно высокие концентрации аллеля I0 обнаруживаются в популяциях аборигенов Австралии и Полинезии, индейцев Америки, которые практически не поражались чумой.
Частота заболеваемости «натуральной» оспой, тяжесть симптомов, смертность выше у лиц с группой крови А или АВ в сравнении с лицами, имеющими группу крови 0 или В (рис. 12.10). Объяснение состоит в том, что у людей первых двух групп отсутствуют антитела, частично нейтрализующие оспенный антиген А. Лица с группой крови 0 в среднем имеют возможность прожить дольше, однако для них выше вероятность заболеть язвенной болезнью.
Таблица 12.3. Примеры аллелей, имеющих адаптивное значение
Аллели и генотипы |
Географическая распространенность |
Адаптивное значение |
Группы крови системы АВ0, аллель В
Аллель А
Трансферрины — белки, связывающие железо, аллель Tf DI
Кислая фосфатаза эритроцитов, аллель Acpr
Аллель АсР и группа крови АВ
Аллель АсР и группа крови А или аллель АсР и группа крови АВ
Сухая ушная сера, аллель d
|
Повсеместно, чаще в Азии
Повсеместно
Высокая частота в зоне тропического пояса
Высокая частота у бушменов и негроидов Центральной Африки
Высокая частота на о.Новая Гвинея
Высокая частота у жителей Чукотки и Аляски
Высокая частота в Дальневосточном регионе
|
Относительная устойчивость к чуме
Относительная устойчивость к язве желудка и двенадцатиперстной кишки
Резистентность ко многим инфекционным заболеваниям
Высокая активность фермента при повышениях температуры
Устойчивость к тропи ческой малярии Легкость адаптации в условиях холодного климата
При генотипе dd низки уровень холестерина и концентрация липидов в крови, высокая концентрация лизоцима в ушной сере |
Вместе с тем для популяций из одного географического района, но изолированных в репродуктивном отношении, причиной различий в концентрации аллелей АВО мог быть дрейф генов. Так, частота группы крови А достигает у индейцев племени черноногих 80%, а у индейцев из штата Юта — 2%.
В основе стойкого сохранения в популяции людей одновременно нескольких аллелей одного гена лежит, как правило, отбор в пользу гетерозигот, который ведет к состоянию балансированного полиморфизма. Классическим примером такой ситуации является распространение аллелей гемоглобинов S, С, и Е в очагах тропической малярии.
Выше приведены примеры полиморфизма по конкретным локусам, который объясняется действием известного фактора отбора. В естественных условиях в силу воздействия на фенотипы организмов комплекса факторов отбор осуществляется по многим направлениям. В результате формируются генофонды, сбалансированные по набору и частотам аллелей, обеспечивающие в данных условиях достаточную выживаемость популяций. Это справедливо и для популяций людей. Так, люди с группой крови 0 более восприимчивы к чуме, чем люди с группой В. Туберкулез легких у них лечится с большим трудом, чем у лиц с группой крови А. Вместе с тем лечение больных сифилисом людей с группой крови 0 быстрее вызывает переход болезни в неактивную стадию. Для лиц с группой крови 0 вероятность заболеть раком желудка, раком шейки матки, ревматизмом, ишемической болезнью сердца, холециститом, жёлчно-каменной болезнью примерно на 20% ниже, чем для лиц с группой А.
Генетический полиморфизм по многим локусам мог быть унаследован людьми от предков на досапиентной стадии развития. Полиморфизм по таким системам групп крови, как АВ0 и Rh, обнаружен у человекообразных обезьян. Факторы отбора, действие которых создавало современную картину распределения аллелей в популяции людей, для подавляющего большинства локусов точно не установлены. Примеры, рассмотренные выше, указывают на их экологическую природу.
Учитывая слабую техническую оснащенность, плохие экономические и гигиенические условия жизни основной массы населения планеты на протяжении значительной части истории человечества, можно представить, какую большую роль играли возбудители особо опасных инфекций, паразитарных заболеваний, туберкулеза. В этих условиях наследственный полиморфизм способствовал расселению людей, обусловливая удовлетворительную жизнеспособность в разных экологических ситуациях. Определенный вклад в наблюдаемое распределение аллелей внесли массовые миграции населения и сопутствующая им метизация. В период до Великих географических открытий и начала колониальной экспансии смешение больших контингентов людей разной расовой принадлежности имело место в Восточной Африке, Индии, Средней Азии, Западной Сибири, Алтае-Саянском нагорье, Индокитае. Впоследствии это наблюдалось в Южной и Центральной Америке.
Генетический полиморфизм является основой межпопуляционной и внутрипопуляционной изменчивости людей. Изменчивость проявляется в неравномерном распределении по планете некоторых заболеваний, тяжести их протекания в разных человеческих популяциях, разной степени предрасположенности людей к определенным болезням, индивидуальных особенностях развития патологических процессов, различиях в реакции на лечебное воздействие. Наследственное разнообразие долго было препятствием успешному переливанию крови. В настоящее время оно же создает большие трудности в решении проблемы пересадок тканей и органов.
Генетический груз в популяциях людей
Так же как и в популяциях других организмов, наследственное разнообразие снижает реальную приспособленность популяций людей. Бремя генетического груза человечества можно оценить, введя понятие летальных эквивалентов. Считают, что число их в пересчете на гамету колеблется от 1,5 до 2,5 или от 3 до 5 на зиготу. Это означает, что то количество неблагоприятных аллелей, которое имеется в генотипе каждого человека, по своему суммарному вредному действию эквивалентно действию 3—5 рецессивных аллелей, приводящих в гомозиготном состоянии к смерти индивидуума до наступления репродуктивного возраста.
При наличии неблагоприятных аллелей и их сочетаний примерно половина зигот, образующихся в каждом поколении людей, в биологическом плане несостоятельна. Такие зиготы не участвуют в передаче генов следующему поколению. Около 15% зачатых организмов гибнет до рождения, 3 — при рождении, 2 — непосредственно после рождения, 3 — умирает, не достигнув половой зрелости, 20 — не вступают в брак, 10% браков бездетны.
Неблагоприятные последствия генетического груза в виде рецессивных аллелей, если они не приводят к гибели организма, проявляются в снижении ряда важных показателей состояния индивидуума, в частности его умственных способностей. Исследования, проведенные на популяции арабов в Израиле, для которой характерна высокая частота близкородственных браков (34% между двоюродными и 4% между дважды двоюродными сибсами), показали снижение умственных способностей у детей от таких браков.
Исторические перспективы человека в силу его социальной сущности не связаны с генетической информацией, накопленной видом Homo sapiens в ходе эволюции. Тем не менее человечество продолжает «оплачивать» эти перспективы, теряя в каждом поколении часть своих членов из-за их генетической несостоятельности.
113. Влияние мутационного процесса, миграции, изоляции, дрейфа генов на генетическую конституцию людей. Специфика действия естественного отбора в человеческих популяциях.
ВЛИЯНИЕ ЭЛЕМЕНТАРНЫХ ЭВОЛЮЦИОННЫХ ФАКТОРОВ НА ГЕНОФОНДЫ ЧЕЛОВЕЧЕСКИХ ПОПУЛЯЦИЙ
Клеточные и физико-химические механизмы наследственности и изменчивости универсальны для всех живых существ, включая человека. Установлена определенная зависимость жизнеспособности индивидуума от особенностей его генотипа. Большую часть своей истории человечество было совокупностью более или менее изолированных в репродуктивном отношении относительно малочисленных групп. Вплоть до настоящего времени сохраняются изоляты. В отдельные исторические периоды происходили миграции значительных масс людей. Они сопровождались объединением ранее разобщенных групп, освоением новых территорий со своими климато-географическими условиями. В настоящее время миграции населения усилились в связи с ростом численности людей, совершенствованием средств транспорта, неравномерньм развитием экономики. Благодаря отмеченному генофонды популяций людей испытывали ранее и продолжают испытывать действие элементарных эволюционных факторов. Социальность человека вносит в это действие определенную специфику.
Мутационный процесс
Мутационный процесс у человека сходен с таковым у других организмов по всем основным показателям — средней частоте мутирования на локус или геном за поколение, генетико-физиологическим характеристикам мутаций, наличию антимутационных барьеров. Это совпадение неслучайно. Основные характеристики спонтанного мутагенеза формировались на начальных этапах эволюции жизни под действием таких постоянных факторов, как ультрафиолетовое и иные виды излучения, температура, определенная химическая среда.
Хотя оценка частоты возникновения мутаций у людей встречает серьезные трудности, некоторые подходы к получению таких данных имеются. Согласно одному из них, максимальная определяемая вероятность новой мутации составляла 2,24 · 10-5 на один локус в поколении.
В настоящее время давление мутационного процесса на генофонд человечества, по-видимому, усиливается благодаря росту индуцированных мутаций. Их причиной нередко служат факторы, возникающие в связи с производственной деятельностью человека в условиях научно-технической революции, например ионизирующее излучение. Подсчет прироста количества мутаций сверх фоновых значений встречается с теми же трудностями, о которых шла речь выше. Согласно ориентировочным данным, доза в 1 Гр (грей), получаемая при низком уровне радиации мужчинами, индуцирует от 1000 до 2000 мутаций с серьезными фенотипическими последствиями на каждый миллион живых новорожденных. У женщин эта цифра ниже — 900.
Мутагенные факторы индуцируют мутации как в половых, так и в соматических клетках. В последнем случае результат может состоять в повышении частоты определенных заболеваний, прежде всего злокачественных опухолей. В отношении ионизирующих излучений, в частности, это лейкозы. Далее идут рак молочной железы и щитовидной железы.
Популяционные волны
Численность населения планеты за обозримый исторический период в целом возросла. В эпоху неолита (10—6 тыс. лет назад) число людей было равно примерно 5 млн., в период появления городов (4,5—3,5 тыс. лет назад) — 20—40 млн., во времена Римской империи к началу новой эры — 200 млн., к 1600 г. - 500 млн., в 1800 г. -1 млрд., в настоящее время — более 6 млрд. Представляя общую тенденцию в истории человечества, эти данные указывают также на изменение во времени темпов прироста народонаселения. Этот показатель, однако, изменялся в историческом развитии неравномерно.
Моменты ускорения прироста численности людей совпадают с важнейшими достижениями человечества — развитием земледелия примерно 8000 лет назад, началом индустриализации, эрой научно-технической революции. Важным следствием увеличения темпа прироста является изменение плотности населения. Так, в эпоху охотников и собирателей (30—20 тыс. лет назад) она составляла менее 1 человека на 3 км2, в век бронзы и железа (4—3 тыс. лет назад) — 10 человек, в настоящее время — от 4 до 900—1200 на 3 км2. Даже сейчас 50% населения размещаются всего на 5% площади обитаемой суши. Крайне неравномерное распределение людей на Земном шаре имело место на любом этапе развития человечества. Ускорение роста численности при ограниченности заселяемой территории способствует усилению миграций.
На фоне общей тенденции к повышению численности людей имели место отдельные снижения этого показателя. Причиной снижения, отраженного на диаграмме динамики населения планеты, явилась эпидемия чумы с большой смертностью, которая в средние века распространилась на значительные территории. Предполагают, что сокращение численности людей на территории Европы происходило также в начале каменного века. Причина состояла в уничтожении племенами, освоившими технику коллективной охоты, основного источника пищи — крупных травоядных животных. Заметные колебания численности на ограниченных территориях происходили вследствие, например, особо опасных инфекций. Они были закономерными явлениями в жизни целых народов еще в начале текущего столетия.
Периодические колебания численности людей на обширных или ограниченных территориях, изменяя плотность населения и вызывая миграции, влияли на состояние генофондов человеческих популяций.
Изоляция
Человеческое общество длительно развивалось как совокупность изолированных производственных коллективов, внутри которых в основном и совершались браки. Природа изоляционных барьеров между популяциями людей разнообразна. В ранней истории человечества важное место принадлежало, по-видимому, географической изоляции. Специфическими для человеческого общества являются формы изоляции, зависящие от разнообразия культур, экономических укладов, религиозных и морально-этических установок.
Фактор изоляции оказывал влияние на генофонды популяций людей. Длительным проживанием в состоянии относительной культурной и географической изоляции объясняют, например, некоторые антропологические особенности представителей малых народностей: своеобразный рельеф ушной раковины бушменов, большая ширина нижнечелюстного диаметра коряков и ительменов, исключительное развитие бороды айнов. Среди горных таджиков, проживающих в одном районе, выделены группы с разным соотношением индивидуумов по антигенам эритроцитов системы АВО. Причиной различий является изолированность от главных перевальных путей сообщения.
Сохранению высокого уровня генетической изоляции двух популяций, существующих на одной территории, способствуют отличия по физическим признакам или образу жизни. Однако такие барьеры со временем ослабевают. Об этом свидетельствует судьба популяций белых и негров в США и Бразилии. К настоящему времени доля генов от белых составляет у американских негров 25%, а у бразильских — 40%. Между двумя генетически различающимися популяциями, разделенными географически, иногда вклиниваются другие популяции, через которые и происходит обмен генами. В таких случаях наблюдается градиент признака. Так, частота аллеля группы крови В в Европе постепенно повышается с запада на восток. У коренного населения Пиренейского п-ова этот ген практически отсутствует, тогда как в районе Астрахани его частота достигает 30%.
В настоящее время круг возможных браков неуклонно расширяется. Разрушение многовековых изоляционных барьеров — процесс, по-видимому, необратимый.
Генетико-автоматические процессы
Предположительно человеческие популяции в палеолите состояли из нескольких сотен индивидуумов. Всего одно-два столетия тому назад люди жили преимущественно поселениями в 25—35 домов. Вплоть до самого последнего времени количество индивидуумов в отдельных популяциях, непосредственно участвующих в размножении, редко превышало 400—3500 человек. Причины географического, экономического, расового, религиозного, культурного порядка ограничивали брачные связи масштабами определенного района, племени, поселения, секты. Высокая степень репродуктивной изоляции малочисленных человеческих популяций на протяжении многих поколений создавала благоприятные условия для дрейфа генов.
Генетико-автоматические процессы, или дрейф генов, приводят к сглаживанию изменчивости внутри группы и появлению случайных, не связанных с отбором различий между изолятами. Именно это выявили наблюдения за особенностями фенотипов малочисленных групп населения в условиях, например, географической изоляции. Так, среди жителей Памира резус-отрицательные индивидуумы встречаются в 2—3 раза реже, чем в Европе. В большинстве кишлаков такие люди составляют 3—5% популяции. В некоторых изолированных селениях, однако, их насчитывается до 15%, т.е. примерно как в европейской популяции.
В крови человека имеются гаптоглобины, которые связывают свободный гемоглобин после разрушения эритроцитов, чем предотвращают его выведение из организма. Синтез гаптоглобина Нр1-1 контролируется геном Нр1. Частота этого гена у представителей двух соседних племен на Севере Южной Америки составляет 0,205 и 0,895, отличаясь более чем в 4 раза.
Примером действия дрейфа генов в человеческих популяциях служит эффект родоначальника. Он возникает, когда несколько семей порывают с родительской популяцией и создают новую на другой территории. Такая популяция обычно поддерживает высокий уровень брачной изоляции. Это способствует случайному закреплению в ее генофонде одних аллелей и утрате других. В результате частота очень редкого аллеля может стать значительной.
Так, члены секты амишей в округе Ланкастер штата Пенсильвания, насчитывающей к середине девятнадцатого века примерно 8000 человек, почти все произошли от трех супружеских пар, иммигрировавших в Америку в 1770 г. В этом изоляте обнаружено 55 случаев особой формы карликовости с многопалостью, которая наследуется по аутосомно-рецессивному типу. Эта аномалия не зарегистирирована среди амишей штатов Огайо и Индиана. В мировой медицинской литературе описано едва ли 50 таких случаев. Очевидно, среди членов первых трех семей, основавших популяцию, находился носитель соответствующего рецессивного мутантного аллеля — «родоначальник» соответствующего фенотипа.
В XVIII в. из Германии в США иммигрировало 27 семей, основавших в штате Пенсильвания секту дункеров. За 200-летний период существования в условиях сильной брачной изоляции генофонд популяции дункеров изменился в сравнении с генофондом населения Рейнской области Германии, из которой они произошли. При этом степень различий во времени увеличивалась. У лиц в возрасте 55 лет и выше частоты аллелей системы групп крови MN ближе к цифрам, типичным для населения Рейнской области, чем у лиц в возрасте 28—55 лет. В возрастной группе 3—27 лет сдвиг достигает еще больших значений (табл. 12.1).
Таблица 12.1. Прогрессивное изменение концентрации аллелей системы групп крови MN в популяции дункеров
Возраст членов изолята, лет |
Концентрация аллеля LM |
LN |
Более 55 От 28 до 55 От 3 до 27 |
0,55 0,66 0,735 |
0,45 0,34 0,265 |
Рост среди дункеров лиц с группой крови М и снижение — с группой крови N нельзя объяснить действием отбора, так как направление изменений не совпадает с таковым в целом для населения штата Пенсильвания. В пользу дрейфа генов говорит также то, что в генофонде американских дункеров увеличилась концентрация аллелей, контролирующих развитие заведомо биологически нейтральных признаков, например оволосения средней фаланги пальцев, способности отставлять большой палец кисти.
На протяжении большей части истории человечества дрейф генов оказывал влияние на генофонды популяций людей. Так, многие особенности узкоместных типов в пределах арктической, байкальской, центрально-азиатской, уральской групп населения Сибири являются, по-видимому, результатом генетико-автоматических процессов в условиях изоляции малочисленных коллективов. Эти процессы, однако, не имели решающего значения в эволюции человека.
Последствия дрейфа генов, представляющие интерес для медицины, заключаются в неравномерном распределении по группам населения Земного шара некоторых наследственных заболеваний. Так, изоляцией и дрейфом генов объясняется, по-видимому, относительно высокая частота церебромакулярной дегенерации в Квебеке и Ньюфаундленде, детского цестиноза во Франции, алкаптонурии в Чехии, одного из типов порфирии среди европеоидного населения в Южной Америке, адреногенитального синдрома у эскимосов. Эти же факторы могли быть причиной низкой частоты фенилкетонурии у финнов и евреев-ашкенази.
Изменение генетического состава популяции вследствие генетико-автоматических процессов приводит к гомозиготизации индивидуумов. При этом чаще фенотипические последствия оказываются неблагоприятными. Вместе с тем следует помнить, что возможно образование и благоприятных комбинаций аллелей. В качестве примера рассмотрим родословные Тутанхамона и Клеопатры VII, в которых близкородственные браки были правилом на протяжении многих поколений.
Тутанхамон умер в возрасте 18 лет. Анализ его изображения в детском возрасте и подписи к этому изображению позволяют предположить, что он страдал генетическим заболеванием — целиакией, которая проявляется в изменении слизистой оболочки кишечника, исключающем всасывание клейковины. Тутанхамон родился от брака Аменофиса III и Синтамоне, которая была дочерью Аменофиса III. Таким образом, мать фараона была его сводной сестрой. В могильном склепе Тутанхамона обнаружены мумии двух, по всей видимости мертворожденных, детей от брака с Анкесенамон, его племянницей. Первая жена фараона была или его сестрой, или дочерью. Брат Тутанхамона Аменофис IV предположительно страдал болезнью Фрелиха и умер в 25—26 лет. Его дети от браков с Нефертити и Анкесенамон (его дочерью) были бесплодны. С другой стороны, известная своим умом и красотой Клеопатра VII была рождена в браке сына Птоломея Х и его родной сестры, которому предшествовали кровнородственные браки на протяжении по крайней мере шести поколений.
Естественный отбор
В процессе видообразования естественный отбор переводит случайную индивидуальную изменчивость в биологически полезную групповую — популяционную, видовую. Стабилизирующая его форма сохраняет «удачные» комбинации аллелей от предшествующих этапов эволюции. Отбор поддерживает также состояние генетического полиморфизма. Смена биологических факторов исторического развития социальными привела к тому, что в человеческих популяциях отбор утратил функцию видообразования. За ним сохранились функции стабилизации генофонда и поддержания наследственного разнообразия.
В пользу действия стабилизирующей формы естественного отбора говорит, например, большая смертность среди недоношенных и переношенных новорожденных по сравнению с доношенными. Направление отбора среди таких детей зависит, по-видимому, от снижения общей жизнеспособности. Отрицательный отбор по одному локусу можно проиллюстрировать на примере системы групп крови «резус» (Rh).
Около 85% населения Европы имеет в эритроцитах антиген Rh и образует группу Rh-положительных индивидуумов. Остальные люди из европейской популяции лишены этого антигена и являются Rh-отрицательными. Синтез антигена Rh контролируется доминантным аллелем D и происходит у лиц с генотипами DD и Dd. Резус-отрицательные люди являются рецессивными гомозиготами (dd). При беременности Rh-отрицательной женщины (dd) Rh-положительным плодом (мужчина DD или Dd, плод Dd) при нарушении целостности плаценты в родах Rh-положительные эритроциты плода проникают в организм матери и иммунизируют его (рис. 12.8). При последующей беременности Rh-положительным плодом (Dd) анти-Rh-антитела проникают через плаценту в организм плода и разрушают его эритроциты. Развивается гемолитическая болезнь новорожденного. Ведущим ее симптомом является тяжелая анемия.
В настоящее время медицина располагает способами борьбы с этой формой патологии в виде быстрого переливания младенцу Rh-отрицательной крови или введения анти-Rh-антител для предотвращения иммунизации матери. В отсутствие медицинской помощи новорожденный с гемолитической болезнью нередко погибал. При Rh-отрица-тельном фенотипе матери Rh-положительный плод всегда гетерозиготен (Dd). Это означает, что со смертью такого индивидуума из генофонда популяции, к которой он принадлежит, удаляется равное количество доминантных и рецессивных аллелей локуса «резус». Отбор в данном случае направлен против гетерозигот. При неравенстве исходных частот удаляемых из генофонда аллелей такой отбор приводит к постепенному снижению доли более редкого из них. В европейской популяции таковым является рецессивный аллель d. Подсчитано, что снижение его доли с 15 до 1% путем отбора против гетерозигот потребует 600 поколений, или около 15 000 лет.
Под действием отбора находятся аллели, контролирующие синтез антигенов системы групп крови АВ0. Об этом свидетельствует снижение против ожидаемого количества детей в АВ0-несовместимых браках женщин 0-группы с мужчинами А, В или АВ-групп.
Особенность отбора по локусам группоспецифичных антигенов эритроцитов, таких, как «резус» или АВО, заключается в том, что он действует на уровне зиготы или ранних стадий эмбриогенеза. Замечено, что в браках, где мать и отец несовместимы по фактору «резус», число детей с гемолитической болезнью меньше ожидаемого. Так, среди белого населения США Rh-отрицательные люди составляют примерно 16%. В такой популяции частота аллеля d равна 40%, а вероятность развития гемолитической болезни при случайном подборе брачных пар — 9%. На самом деле частота этой болезни составляет 1:150—1:200. Одним из факторов снижения является одновременная несовместимость родителей по локусу АВ0. В этом случае Rh-положительные эритроциты плода, попавшие в организм матери, разрушаются и иммунизации не происходит.
Отрицательный отбор действует в большинстве популяций людей по аллелям аномальных гемоглобинов. Его особая жесткость обусловливается тем, что он направлен против гомозигот. Ребенок, умирающий, например, от серповидно-клеточной анемии, является гомозиготным по аллелю S. Каждая такая смерть устраняет из генофонда популяции аллели одного вида. Это приводит к сравнительно быстрому снижению изменчивости по соответствующему локусу. Во многих популяциях людей частота аллелей аномальных гемоглобинов, в том числе и S, не превышает 1%.
Высокая частота аллелей таких аномальных гемоглобинов, как S, С, D, Е, в некоторых районах планеты иллюстрирует действие естественного отбора по поддержанию в человеческих популяциях состояния балансированного генетического полиморфизма. Отрицательный отбор в отношении аллеля S перекрывается мощным положительным отбором гетерозигот HbAHbS благодаря высокой жизнеспособности последних в очагах тропической малярии.
Исследования в Уганде показали, что количество возбудителей в 1 мл крови зависит от генотипа ребенка и составляет до 10 000 у HbSHbS, до 160 000 у HbAHbS и до 800 000 у НbAHbА. В Северной Греции обследовали 48 семей, в которых наблюдались и серповидно-клеточность, и малярия. Среди братьев и сестер, больных серповидно-клеточной анемией, болело малярией 16 из 25 с генотипами HbAHbA и 1 из 23 с генотипами HbAHbS.
Таким образом, нормальные люди болели в 13 раз чаще, чем индивидуумы с аномальным генотипом. Устранение фактора контротбора приводит к снижению частоты аллеля серповидно-клеточности. Этой причиной, действующей на протяжении уже нескольких столетий наряду с метисацией, объясняют относительно низкую частоту гетерозигот HbAHbS среди североамериканских негров (8—9%) в сравнении с африканскими (около 20%).
В приведенных примерах действию отрицательного отбора, снижающего в генофондах некоторых популяций людей концентрацию определенных аллелей, противостоят контротборы, которые поддерживают частоту этих аллелей на достаточно высоком уровне. Результатом наложения многочисленных и разнонаправленных векторов отбора является формирование и поддерживание генофондов популяций в состоянии, обеспечивающем возникновение в каждом поколении генотипов достаточной приспособленности с учетом местных условий. Благодаря социально-экономическим преобразованиям, успехам лечебной и особенно профилактической медицины влияние отбора на генетический состав популяций людей прогрессивно снижается.
112. Системы браков. Роль системы браков в распределении аллелей в популяции. Кровнородственные и ассортативные браки.
На генетическую структуру популяции человека (частота аллелей, частота генотипов, величина генетического груза, частота наследственной патологии) оказывает влияние система браков, преобладающая в данной популяции.
Системы браков по критерию выборочности

Поддерживающие равнвесие генотипов Изменяющие соотношение
В популяции генотипов в популяции
популяции генотипов в популяции
Ассортативные
Н
 еассортативные
еассортативные
Положительные Отрицательн.
Панмиксные
Панмиксные браки. Панмиксия – свободное скрещивание, когда образование супружеских пар не зависит от генотипических индивидов и является случайной. Панмиксии в человеческих популяциях препятствуют культурные, лингвистические преграды, изоляция расстояний.
Ассортативные браки. Ассортативность – явление, когда выбор партнера неслучаен, осуществляется по ряду критериев предпочтения.
Положительная ассортативность – выбор партнера со схожими признаками, т.е. со сходным генотипом, что приводит к снижению гетерозиготности в потомстве и уменьшению генетического разнообразия популяции.
Примеры положительной ассортативности:
По месту рождения
По возрасту
По образованию
П вероисповеданию
По шизофрении(особенности поведения)
По склонности к алкоголизму и наркомании
Отрицательная ассортативность – выбор партнера с отличающимися признаками, т.е. с различным генотипом, что приводит к увеличению гетерозиготности потомства и увеличению генетического разнообразия популяции.
Примеры:
По цвету волос
По росту
По весу
Система браков по критерию
Родства

Аутбридинг Инбридинг
Поддерживающие равновесие в популяции Изменяющие соотношение генотипов
, т.е. приближается к панмиксным бракам в популяции (вариант положительно
Или отрицательно ассортативным бракам ассортативного брака)
Аутбредные браки. Аутбридинг – брак между партнерами, не состоящими в родстве, что приводит к увеличению гетерозиготности в потомстве и генетического разнообразия в популяции.
Инбредные браки. Инбридинг – брак между партнерами, состоящими в родстве, что приводит к увеличению уровня гомозиготности в потомстве и уменьшению генетического разнообразия популяции.
По степени родства инбредные браки подразделяют на:
Инцест – запретный брак (между родственниками 1 степени)
Кровнородственный брак (между родственниками 2 и 3 степени)
Мерой генетических последствий инбридинга служит коэффициент инбридинга – вероятность того, что у особи окажется 2 аллеля (аа), идентичные по происхождению (от общего предка).
У детей (F1) пары родителей вероятность одинаковых аллелей в одном и том же локусе равна 0,5 %. У их детей (F2) эта вероятность равна 0.25 %. Таким образом, при вступлении в брак двоюродных сибсов коэффициент инбридинга равен 1/16 %.
Примеры популяций с инбредными браками:
1) Инцест в человеческих популяциях допускался в редких случаях (династия потомков в государстве инков. В племени эрнадан с Малабарского берега).
2) В странах Ближнего Востока на долю близкородственных браков приходится 20 – 70 %, что приводит к увеличению частоты бесплодия, самопроизвольных выкидышей, врожденных пороков развития, младенческой детской смертности.
3) В некоторых областях Индии с высоким уровнем инбредных браков не отмечается негативного влияния на потомство (инбридинг в течение многих поколений привел к уменьшению вредных аллелей в популяции – гипотеза).
4) В большинстве человеческих популяций допустимый инбридинг – брак между дядей и племянницей или тетей и племянником.
Биологические последствия различных систем браков.
1) Положительная ассортативность в больших популяциях приводит к тенденции увеличения гомозиготности и снижения гетерозиготности, в следствие чего возникает отклонение от панмексии по некоторым наследственным болезням (фенилкетонурия,
Врожденный гепотириоз, муковисцидоз, талассемя и др.).
2) В небольших популяциях даже при случайном выборе партнера отмечается тенденция к инбредности – увеличение q2 (аа) – летальные ген, увеличение 2pq (Аа) по одинаковым аутосомно-рецессивным генам – увеличение частоты АА наследственных болезней (инбредная депрессия) – уменьшения жизнеспособности и плодовитости популяции.
Примеры инбредной депрессии:
а) Частота фенилкетонурии
В неродственных браках 1: 15000
В родственных 1: 7000
б) Частота альбинизма
В неродственных 1: 40000
В родственных 1: 3000
3) В изолятах большая роль принадлежит «эффекту родоначальника» (особенности генотипов людей – основателей изолятов).
Если у основателей имелись летальные гены, то в условиях инбридинга эти гены могут получить широкое распространение – вымирание изолята.
Пример:
а) Секта мармонов-менонитов в США
Несколько тысяч человек из Голландии
Высокая частота карликовой хондродистрофии
б) В ЮАР белое население:
Высокая частота Порфирии
в) Евреи-шикенази
Высокая частота амовратической идиотии.
114. Экология как наука. Предмет, структура, содержание и методы экологии. Экологические факторы и их взаимодействие.
Термин "Экология" предложен Геккелем в 1869 г. Экология ("ойкос" - дом, "логос" - наука) - это наука о взаимодействии организмов с окружающей средой.
Экология — это наука, изучающая закономерности взаимодействия организмов и среды их обитания, законы развития и существования биогеоценозов как комплексов взаимодействующих живых и неживых компонентов в различных участках биосферы.
Объектами экологии являются преимущественно системы выше уровня организмов, т. е. изучение организации и функционирования надорганизменных систем: популяций, биоценозов (сообществ), биогеоценозов (экосистем) и биосферы в целом. Другими словами, главным объектом изучения в экологии являются экосистемы, т. е. единые природные комплексы, образованные живыми организмами и средой обитания.
Задачи экологии меняются в зависимости от изучаемого уровня организации живой материи. Популяционная экология исследует закономерности динамики численности и структуры популяций, а также процессы взаимодействий (конкуренция, хищничество) между популяциями разных видов. В задачи экологии сообществ (биоценологии) входит изучение закономерностей организации различных сообществ, или биоценозов, их структуры и функционирования (круговорот веществ и трансформация энергии в цепях питания).
Главная же теоретическая и практическая задача экологии — раскрыть общие закономерности организации жизни и на этой основе разработать принципы рационального использования природных ресурсов в условиях все возрастающего влияния человека на биосферу.
Взаимодействие человеческого общества и природы стало одной из важнейших проблем современности, поскольку положение, которое складывается в отношениях человека с природой, часто становится критическим: исчерпываются запасы пресной воды и полезных ископаемых (нефти, газа, цветных металлов и др.), ухудшается состояние почв, водного и воздушного бассейнов, происходит опустынивание огромных территорий, усложняется борьба с болезнями и вредителями сельскохозяйственных культур. Антропогенные изменения затронули практически все экосистемы планеты, газовый состав атмосферы, энергетический баланс Земли. Это означает, что деятельность человека вступила в противоречие с природой, в результате чего во многих районах мира нарушилось ее динамическое равновесие.
Для решения этих глобальных проблем и прежде всего проблемы интенсификации и рационального использования, сохранения и воспроизводства ресурсов биосферы экология объединяет в научном поиске усилия ботаников, зоологов и микробиологов, придает эволюционному учению, генетике, биохимии и биофизике их истинную универсальность.
В круг проблем экологии включены также вопросы экологического воспитания и просвещения, морально-этические, философские и даже правовые вопросы. Следовательно, экология становится наукой не только биологической, но и социальной.
Методы экологии подразделяются на полевые (изучение жизни организмов и их сообществ в естественных условиях, т. е. длительное наблюдение в природе с помощью различной аппаратуры) и экспериментальные (эксперименты в стационарных лабораториях, где имеется возможность не только варьировать, но и строго контролировать влияние на живые организмы любых факторов по заданной программе). При этом экологи оперируют не только биологическими, но и современными физическими и химическими методами, используют моделирование биологических явлений, т. е. воспроизведение в искусственных экосистемах различных процессов, происходящих в живой природе. Посредством моделирования можно изучить поведение любой системы с целью оценки возможных последствий применения различных стратегий и методов управления ресурсами, т. е. для экологического прогнозирования.
Для изучения и прогнозирования природных процессов широко используется также метод математического моделирования. Такие модели экосистем строятся на основе многочисленных сведений, накопленных в полевых и лабораторных условиях. При этом правильно построенные математические модели помогают увидеть то, что трудно или невозможно проверить в эксперименте. Однако сама по себе математическая модель не может служить абсолютным доказательством правильности той или иной гипотезы, но она служит одним из путей анализа реальности.
Сочетание полевых и экспериментальных методов исследования позволяет экологу выяснить все аспекты взаимоотношений между живыми организмами и многочисленными факторами окружающей среды, что позволит не только восстановить динамическое равновесие природы, но и управлять экосистемами.
Влияние среды на организмы обычно оценивают через отдельные факторы. Под экологическими факторами понимается любой элемент или условие среды, на которые организмы реагируют приспособительными реакциями или адаптациями. Каждая из сред обитания отличается особенностями воздействия экологических факторов. Все многообразие экологических факторов подразделяют на три группы – абиотические, биотические и антропогенные. Абиотические факторы – компоненты неживой природы. К ним относят: климатические (свет, температура, влажность, ветер, давление и др.), геологические (землетрясения, извержения вулканов, движение ледников, радиоактивное излучение и др.), эдафические или почвенные (плотность, структура, состав почвы), гидрологические (вода, течение, соленость, давление) и другие. Биотические факторы – факторы живой природы. В зависимости от воздействующего организма биотические факторы делят на фитогенные (влияние растений), зоогенные (животных), микробогенные (микроорганизмы), микогенные (грибы). Влияние биотических факторов вызывает рядприспособительных реакций со стороны растений и животных. Антропогенные факторы – факторы человеческой деятельности. Человек вызывает серьезные изменения в биогеоценозах. При этом изменения, производимые им, создают для одних видов благоприятные условия развития, а для других – неблагоприятные. В результате между видами возникают новые численные отношения, перестраиваются пищевые цепи, появляются приспособления, необходимые для существования организмов в измененной среде.
Экологические факторы могут оказывать на организм прямое действие и косвенное, положительное и отрицательное; они обладают различной изменчивостью во времени и в пространстве. Одни из них относительно постоянны (солнечная радиация, соленость океана), другие очень изменчивы (температура и влажность воздуха). Изменения факторов среды могут быть периодическими и непериодическими. Экологические факторы оказывают на живые организмы различные воздействия: ограничивающее (делают невозможным существование в данных условиях), раздражающее (вызывают биохимические и физиологические адаптации), модифицирующее (вызывают морфологические и анатомические изменения организмов), сигнальное (информируют об изменениях других факторов среды). Каждый экологический фактор характеризуется определенными количественными показателями (силой и диапазоном действия). Диапазон определяется как отрезок (амплитуда) в действии фактора, конкретный для каждого организма. Поэтому фактор имеет начальную границу действия, то есть порог включения фактора, и конечную, «верхнюю» границу действия фактора. Благоприятная сила воздействия называется оптимальной зоной экологического фактора или оптимумом. Угнетающее действие (максимальное или минимальное) называется зоной пессимума. Экологические факторы обычно действуют не поодиночке, а целым комплексом. При этом действие одного какого-либо фактора зависит от уровня действия других. Необходимо отметить, что совокупность факторов действует сильнее всего на те фазы развития организмов, которые имеют наименьшую экологическую валентность – минимальную способность к приспособлению.