

Монотопный белок, а-спиральный домен которого пронизывает мембрану.
2. Политопный белок,пронизывающий мембрану несколько раз.
глобулярные белки.
Молекулярный вес 10 – 500 кДа
Много гидрофобных аминокислотных остатков (23-24).
Достаточное количество полярных остатков.
Функции: транспортная (ионные каналы), активный насосный механизм,формирование межклеточных контактов
Полуинтегральные белки
Глобулярные
Экспонируются только на одной поверхности мембраны.
В этом случае молекула белка состоит из двух участков – гидрофобного «якоря» и гидрофильного функционального участка.
Функции
специфические рецепторы,транспортная (белки-переносчики),ферментные белки,переносчики электронов
Периферические белки
Глобулярные и фибриллярные
взаимодействуют только с полярными компонентами – головками фосфолипидов и немембранными доменами интегральных белков. На поверхности мембран периферические белки удерживаются за счёт электростатических сил и солевых мостиков
На наружной поверхности - ламинин и фибронектин, на внутренней - спектрин и альфа-актинин.
Функции: Рецепторные,Адгезионные ,Белки цитоскелета ,Белки системы вторичных посредников,Антитела,Ферменты
Асимметрия белкового состава
Структурная асимметрия тесно связана с функциональной.
На внешней стороне мембраны находятся рецепторные и маркерные молекулы. Внутренняя сторона обеспечивает выполнение исполнительских функций, на ней расположены ферменты, белки, связанные с цитоскелетом и т.д.
Углеводные компоненты
Углеводные остатки различной длины (моно- и олигосахариды) всегда связаны ковалентными связями либо с липидами, либо с белками.
Углеводные цепи плазматической мембраны экспонированы во внеклеточную среду, располагаясь над плоскостью липидного бислоя.
Углеводные компоненты обнаруживаются и в составе мембран внутриклеточных органоидов. В этом случае они экспонированы во внутренний просвет органоида.
Углеводный состав
Среди мономеров в составе углеводных цепей - галактоза и манноза,
реже – ксилоза и глюкоза, и сиаловая кислота. Терминальным мономером является фукоза или сиаловая кислота.
В молекулах гликолипидов моно- и олигосахариды присоединены взамен фосфатных групп.
Функции углеводов мембран
Углеводные цепи этих соединений формируют основную часть гликокаликса эукариотических клеток
Большая часть мембранных рецепторов – гликопротеиды или ганглиозиды, углеводная составляющая которых играет важную роль в реализации трансмембранной передачи сигнала.
Гликопротеидами являются и отдельные компоненты ионных насосов, например, малая субъединица Na+/K+-АТФазы.
Антигены групп крови АВ0, локализованные на плазматической мембране эритроцитов, представляют собой углеводные компоненты гликопротеидов.
Неорганические компоненты мембран
Вода. 30% веса мембраны. Само существование липидного бислоя возможно как раз по причине водного окружения мембран.
Ca2+, Mg2+, K+ и Na+. связываются с участками кислых фосфолипидов, а также белков, включающих гидрофильные кислые домены. Они оказывают влияние на структурно-функциональныые характеристики мембран. Это влияние реализуется через изменение конформационного состояния белков, статического заряда поверхностей мембран и т.д. Обеспечивают ионный гомеостаз
Некоторые мембранные белки содержат ионы металлов.
Функции мембраны
Транспортная
Избирательная проницаемость
Коммуникационная
Межклеточная адгезия
Межклеточные контакты
Ферментативная
Мембраны несут специфические рецепторы, связывающие биологически активные соединения
Функции
благодаря рецепторной и ферментативной активности мембран осуществляется трансмембранная передача сигналов
на мембранах находятся специфические молекулярные маркеры, позволяющие опознавать данный тип мембранного органоида (в случае плазмолеммы – тип клетки)
мембраны обеспечивают сопряжение процессов окисления и фоторецепции с синтезом макроэргических соединений
Транспорт: Пассивный транспорт, Диффузия, Облегченная диффузия, Осмос Активный транспорт Экзо - или эндоцитоз.
Пассивный транспорт: диффузия
Движение молекул по градиенту концентрации
Холестерин, бензол
Небольшие неполярные молекулы О2 и N2
Небольшие полярные Н2О, С О2 , этанол
Облегченная диффузия Ионный канал Ионные каналы (натриевые, кальциевые, аквапорины)
Потенциалзависимые, механочувствительные, рецепторзависимые, кальций-зависимые)
Лиганд-зависимые ионные каналы
Эти каналы открываются, когда медиатор, связываясь с их наружными сторонами, меняет их конформацию. Открываясь, они впускают ионы, изменяя этим мембранный потенциал. Лиганд-зависимые каналы почти нечувствительны к изменению мембранного потенциала.
Облегченная диффузия Белки-переносчики
Белки-переносчики (глюкоза, аминокислоты, анионообменники – обмен Cl- на бикарбонат, фосфаты, обмен Na+ на Н+, норадреналин)
По градиенту концентрации без затраты энергии, м.б. сочетанным, специфичен
Активный транспорт
Энергозависимый трансмембранный перенос против электрохимического градиента, происходит при участии АТФаз
Na+К+-, H+,K+- и Ca2+- АТФазы
Системы антипорта
или контр-транспорта – это система вторично активного транспорта, функционирующая на основе переносчика обменника, обеспечивающего выведение из клетки транспортирующего вещества против его концентрационного градиента в обмен на сопряженный, пассивно поступающий в клетки поток ионов Na+. Движущей силой является потенциальная энергия концентрационного градиента ионов Na+.
Антипортная система клеток проксимальных отделов нефрона, обменивающая Nа+ на Н+. Из мочи к клетки проксимальных отделов нефрона выделяется Nа+, взамен выводится Н+. Система не совершает электрическую работу, поэтому не надо тратить энергию.
Везикулярный перенос Крупные молекулы биополимеров практически не проникают сквозь плазмолемму. В ряде случаев макромолекулы и даже их агрегаты, а часто и крупные частицы попадают внутрь клетки в результате процесса эндоцитоза . Эндоцитоз формально разделяют на фагоцитоз (захват и поглощение клеткой крупных частиц, например бактерий или фрагментов других клеток) и пиноцитоз (захват отдельных молекул и макромолекулярных соединений). Макромолекулы такие как белки, нуклеиновые кислоты, полисахариды,липопротеидные комплексы и другие сквозь клеточные мембраны непроходят, в противовес тому как транспортируются ионы и мономеры.Транспорт микромолекул, их комплексов, частиц внутрь клетки и из нее происходит совершенно иным путем - посредством везикулярногопереноса. Т.е макромолекулы, не могут попадать в клетку сквозьплазматическую мембрану. В клетку они попадают заключенными внутри вакуолей или везикул. Такой везикулярный перенос делят надва вида: экзоцитоз - вынос из клетки макромолекулярных продуктов, и эндоцитоз – поглощение клеткой макромолекул.
Эндоцитоз Эндоцитоз начинается с сорбции на поверхности плазмолеммы поглощаемых веществ. Связывание их с плазмолеммой определяется наличием на ее поверхности рецепторных молекул. После сорбции веществ на поверхности плазмолемма начинает образовывать сначала небольшие впячивания внутрь клетки. Они могут иметь вид еще не замкнутых округлых пузырьков или представлять собой глубокие инвагинации, впячивания внутрь клетки. Затем такие локальные впячивания отшнуровываются от плазмолеммы и в виде пузырьков свободно располагаются под ней.
В такую первичную вакуоль, или в эндосому, могут попадать любые
биополимеры, макромолекулярные комплексы, части клеток или даже целые клетки, где затем и распадаются, деполимеризуются до мономеров,
которые путем трансмембранного переноса попадают в гиалоплазму.
Клетка эпителия тонкого кишечника образует до 1000 пиносом в секунду, а макрофаги образуют около 125 пиносом в минуту.
Размер пиносом составляет 60-130 нм
Фагоцитоз захват клеткой крупных частицх, встречается среди клеток животных, как дноклеточных (амебы, некоторые хищные инфузории), так и в специализированных клетках многоклеточных
животных. Специализированные клетки- фагоциты-характерны для беспозвоночных животных
(амебоциты крови или полостной жидкости), и для позвоночных (нейтрофилы и макрофаги).
Эндоцитоз может быть неспецифическим и специфическим, опосредуемым рецепторами
(рецепторным). Неспецифический эндоцитоз протекает автоматически и часто приводит к захвату и поглощению совершенно чуждых или безразличных для клетки веществ( сажа или красители).
Рецепторный эндоцитоз
Специальные рецепторы плазматической мембраны, диффузно расположенные на поверхности различных клеток, узнают вещество, и образуют специфический комплекс рецептор-лиганд. Вслед за этим такой комплекс перемещается в зону окаймленных ямок и интернализуется - окружается мембраной и погружается вглубь цитоплазмы. Со стороны цитоплазмы плазматическая мембрана покрыта тонким (около 20 нм) волокнистым слоем, окаймляет небольшие ямки. Занимают около 2% клеточной поверхности. Состоит в основном из белка клатрина. Клатриновый слой одевает весь периметр отделяющихся первичных эндоцитозных вакуолей - окаймленных пузырьков. Эти белки связываются с интегральными белками-рецепторами со стороны цитоплазмы и образуют одевающий слой по периметру возникающей пиносомы, - “окаймленного” пузырька. Затем окаймленный пузырек отделится от плазмолеммы и переносится вглубь цитоплазмы, клатриновый слой распадается. После потери клатринового слоя эндосомы начинают сливаться друг с другом.
Основное биологическое значение эндоцитоза
получение строительных блоков за счет внутриклеточного переваривания, котороеосуществляется на втором этапе эндоцитоза после слияния первичной эндосомы с лизосомой.
Экзоцитоз- процесс, обратный эндоцитозу. Внутриклеточные продукты, заключенные в вакуоли и отграниченные от гиалоплазмы мембраной, подходят к плазматической мембране. В местах их контактов плазматическая мембрана и мембрана вакуоли сливаются, и пузырек опустошается в окружающую среду. С экзоцитозом связано выделение синтезированных в клетке разнообразных веществ. Секретирующие, выделяющие вещества во внешнюю среду, клетки могут вырабатывать и выбрасывать низкомолекулярные соединения (ацетилхолин, биогенные амины и др.), а также в большинстве случаев макромолекулы (пептиды, белки, липопротеиды, пептидогликаны и др.). Экзоцитоз происходит в ответ на внешний сигнал (нервный импульс, гормоны, медиаторы . Значение экзоцтоза
секреция веществ для других клеток (секреция молока, пищеварительных соков,гормонов и др.).
секреция для собственных нужд. (рост плазматической мембраны, элементы гликокаликса)
экзоцитоз гидролитических ферментов для пристеночного пищеварения
Адгезия клеток. Одна из важнейших функций плазматических мембран - обеспечение сцепления клеток друг с другом. Основой для такого типа контакта может быть органическая соль кальция, образующая связи с карбоксильными группами белков и липидов. Взаимодействие клеток существенно зависит от величины поверхностного заряда. Отрицательный заряд на поверхности клеток вызывает их взаимное отталкивание. Если смешать клетки зародышей разных видов, то их последующая адгезия будет связана не с видовой специфичностью, а с тканевой принадлежностью. Способность к адгезии у всех клеток различна: клетки злокачественных опухолей обладают более низкой способностью к адгезии, чем нормальные, что объясняется пониженным содержанием Са2+ в раковых клетках и соответственно их высоким поверхностным зарядом.
Надмембранные структуры
Гликокаликс
представляет собой внешний по отношению к мембране слой, содержащий полисахаридные цепочки мембранных интегральных белков - гликопротеидов. Эти цепочки содержат такие углеводы как манноза, глюкоза, n-ацетилглюкозамин, сиаловая кислота и др. Такие углеводные гетерополимеры образуют ветвящиеся цепочки, между которыми могут располагаться выделенные из клетки гликолипиды и протеогликаны. Слой гликокаликса сильно обводнен, имеет желеподобную консистенцию, что значительно снижает в этой зоне скорость диффузии различных веществ. Здесь располагаются выделенные клеткой гидролитические ферменты, участвующие во внеклеточном расщеплении полимеров (внеклеточное пищеварение) до мономерных молекул, которые затем транспортируются в цитоплазму через плазматическую мембрану.
В электронном микроскопе, при специальных методах контрастирования полисахаридов, гликокаликс имеет вид рыхлого волокнистого слоя, толщиной 3-4 нм, покрывающего всю поверхность клетки. Особенно хорошо гликокаликс выражен в щеточной каемке клеток всасывающего эпителия кишечника (энтероциты) .
Состав: Толщина – 45-55 нм. олигосахариды,периферические белки, гликопротеиды, гликолипиды, сиаловая кислота
Строение: Ячеистая структура сети гликокаликса обусловлена наличием анионно-катионных подвижных мостиков между нитями. Размер ячеек сети гликокаликса уменьшается по направлению к плазмалемме.
Функции: Он предназначен для восприятия, распознавания, передачи, модуляции сигналов клетки в виде вещества, энергии, информации
Выступающие разветвленные части гликокаликса обеспечивают:
Межклеточное узнавание
индивидуальность клетки;
иммунологическую индивидуальность клетки (роль антигенов);
Межклеточные взаимодействия соединение клеток при образовании тканей.
Межклеточное узнавание Это специфическое взаимодействие клетки с другой клеткой или с внеклеточным матриксом.
После узнавания
Прекращение миграции клеток
Адгезия клеток
Образование контактов
Образование клеточных ансамблей
Взаимодействие клеток между собой и клетками других структур
Семейство кадгеринов – трансмембранные гликопротеины
Иммуноглобулины.
интегрины (CAD/CAM молекулы).
распознавание клетками друг друга при формировании тканевых структур, взаимодействие клеток с компонентами межклеточного вещества и прикрепление к ним.
Пристеночное пищеварение
В гликокаликсе энтероцитов начинается примембранное пищеварение которое сопрягает процессы полостного пищеварения и всасывания питательных веществ в тонкой кишке.
Индивидуализация
Углеводный компонент гликокаликса играет важную роль в индивидуализации разных типов клеток. Презентация– лимфоциты которые в зависимости от дифференцировки на них антигенов подразделяются на несколько типов – В-, Т-лимфоциты и НК-клетки
Создание поверхностного заряда
У эритроцитов млекопитающих развит углеводный компонент интегрального гликопротеина – гликофорина, который необходим для создания отрицательного заряда на поверхности эритроцитов
Субмембранный слой(Кортикальный)
Кортикальный (cortex -кора) слой цитоплазмы,
лежащий в тесном контакте с мембраной. Основным фибриллярным компонентом кортикального слоя является сеть актиновых микрофибрилл.
Здесь же располагается ряд вспомогательных белков, необходимых для движения участков цитоплазмы.
Компоненты цитоскелета
1.микротрубочки 22±2нм
2. микрофиламенты 5-7нм (актин,миозин, актинин, тропомиозин)
3. промежуточные филаменты -10 нм (прекератин, виментин, десмин) –
Цитоскелет эндотелиоцитов представлен комплексом механохимических белков (тубулин, актин, виментин и др.), обеспечивающих поддержание формы, подвижность клеток. Состоит из варьирующих по распределению и ориентации микротрубочек, микрофиламентов, якорных фибрилл, соединяющих фибриллярные элементы цитоплазмы с цитолеммой и ядерной оболочкой.
Одномембранные органеллы клетки
Мембранная система клетки
Эндоплазматический ретикулум Пластинчатый комплексЛизосомы Пероксисомы Везикулы и др.
Отличительной чертой мембранной системы является то, что синтезированные полимеры и продукты их превращений отделены от собственно цитоплазмы и становятся изолированными от цитозольных ферментов. Такое разобщение очень важно для одновременного протекания в клетке многих синтетических процессов.
Открытие ЭПР
Открыт в 1945 г. К. Портером при изучении фибробластов цыплят в электронном микроскопе. В световом микроскопе в фибрибластах послефиксации и окраски видно, что периферия клеток (эктоплазма) окрашивается слабо, в то время как центральная часть клеток (эндоплазма) хорошо воспринимает красители. Портер увидел в электронном микроскопе, что зона эндоплазмы заполнена большим числом мелких вакуолей и каналов, соединяющихся друг с другом и образующих что-то наподобие рыхлой сети (ретикулум). Было видно, что стопки этих вакуолей и канальцев ограничены тонкими мембранами.
Эндоплазматический ретикулум
1. является местом образования всех мембран клетки, синтезируя и встраивая в существующие мембраны специфические для них белки и липиды). Мембраны ПС разграничивают пространство клетки
2. Внутри канальцев ПС осуществляется транспорт и накопление вновь синтезированных веществ, а также детоксикация метаболитов.
3. синтез веществ, продуцируемых в большом количестве клетками на экспорт
Гладкий ЭПР
Гладкая ЭПС отличается от гранулярной отсутствием связанных рибосом.
а) Обычно в её состав входят соединяющиеся друг с другом небольшие вакуоли и трубочки (1).
1.синтез, транспорт и накопление мембранных и немембранных липидов.
2.Гладкая ПС хорошо развита в клетках коры надпочечников, где специализируется на синтезе стероидных гормонов,
3. В клетках печени, обеспечивая детоксикацию вредных метаболитов, лекарственных веществ
4. В секреторных клетках некоторых растений (синтез терпеноидов).
5.Особенно сильно гладкая ПС развита в мышечных волокнах скелетной мускулатуры, где она формирует L-систему, которая обеспечивает внутриклеточный транспорт ионов кальция с помощью встроенных в мембраны кальциевых насосов.
6. Синтез углеводов (гликоген)
Детоксикация
Одной из наиболее важной функцией гладкого ЭР является детоксикация (при помощи оксидаз гепатоцитов) как продуктов клеточного метаболизма, так и поступающих извне веществ, в том числе этанола и барбитуратов. С участием гладкого ЭР вещества конвертируются в водорастворимые соединения, что способствует их выведению из организма. Для эффективной детоксикации гладкий ЭР в течение нескольких дней может удвоить общую площадь своей поверхности.
Системы:
1. Цитохром P450–зависимая монооксигеназная система локализована в гладком ЭР и состоит из цитохрома P450; НАДФ–P450 редуктазы, которая переносит электроны от редуцируемой формы НАДФ к цитохрому P450; фосфатидилхолина. Активность этой системы наиболее высока в печени, а также в коже, лёгких, слизистой оболочке пищеварительного тракта.
2. Флавин-содержащая монооксигеназная система также характерна для гладкого ЭР гепатоцитов. Окисляет различные амины, в том числе и никотин.
3. Пероксидаза-зависимое коокисление катализируется простагландин H синтазой (ферментом, участвующем в метаболизме арахидоновой кислоты). Фермент локализован в гладком ЭР; активность особенно высока в семенных пузырьках, почке и мочевом пузыре. Простагландин-H синтаза метаболизирует 2-нафтиламин — соединение, присутствующее в синтетических препаратах и повышающее риск развития рака мочевого пузыря.
В ходе всех реакций детоксикации попутно могут образовываться токсичные активные формы кислорода. Для борьбы с активными формами кислорода клетки обладают рядом защитных механизмов и ферментов, таких, как супероксид дисмутаза, каталаза, глутатион редуктаза. В качестве эндогенных антиоксидантов могут выступать витамины C, E, b-каротин.
Синтез стероидных гормонов
В стероидпродуцирующих клетках (коры надпочечников, половых желёз) гладкий ЭР служит для метаболизма стероидов и образования конечных форм стероидных гормонов с участием митохондрий. Сначала в митохондриях от молекулы холестерина отсекается боковая цепь, после чего полученный продукт поступает в гладкий ЭР, где происходит его дальнейшая модификация. Образованное вещество вновь попадает в митохондрии, где молекула гормона окончательно формируется. Стероидные гормоны не запасаются в клетке в секреторных гранулах; секреция гормона происходит почти сразу после его синтеза. После высвобождения из клетки в кровоток гормоны связываются со специальными связывающими белкми, транспортирующими стероид к клетке-мишени. Стероидные гормоны оказывают свое влияние на клетку, регулируя транскрипцию генов, в результате чего ответ клетки-мишени возникает спустя несколько часов или дней. В метаболизме стероидных гормонов — помимо эндокринных клеток гонад и надпочечников — участвуют также кератиноциты, гепатоциты, эпителиальные клетки канальцев почек и др.
Депо кальция
Цистерны гладкой эндоплазматической сети многих клеток специализированы для накопления в них Ca2+ путём постоянного откачивания Ca2+ из цитоплазмы,. Подобные депо существуют в скелетной и сердечной мышцах, нейронах, яйцеклетке, эндокринных клетках и т.д.. Различные сигналы (например, гормоны, нейромедиаторы, факторы роста) влияют на функции клеток и их пролиферацию путём изменения концентрации в цитозоле внутриклеточного посредника — Ca2+. Например, условие сокращения мышечных элементов — резкое повышение концентрации Ca2+ в цитозоле. Для этого необходимо постоянно откачивать ионы кальция из цитозоля и накапливать их в специальных депо, образованных Ca2+ -запасающими цистернами гладкой эндоплазматической сети. Внутри цистерн находятся Ca2+ -связывающие белки. В мембрану цистерн — депо Ca2+ встроены Ca2+ -насосы (Ca2+ -АТФаза), постоянно закачивающие Ca2+ внутрь цистерн, и Ca2+ -каналы, через которые происходит выброс Ca2+ из депо при поступлении сигнала.
· Ca2+ -связывающие белки внутри цистерн (кальсеквестрин, кальретикулин и др.) непрочно ассоциированы с Ca2+.Кальсеквестрин — главный Ca2+ -связывающий белок саркоплазматической сети волокон поперечнополосатой мышцы и некоторых ГМК. Одна молекула кальсеквестрина связывает приблизительно 50 ионов Ca2+ . Кальретикулин присутствует в саркоплазматической сети большинства ГМК и в эндоплазматическом ретикулуме немышечных клеток.
Гранулярный ЭПР
На ультратонких срезах гранулярный ЭР представлен замкнутыми мембранами, которые образуют на сечениях вытянутые мешки, цистерны или же имеют вид узких каналов. Ширина полостей цистерн может очень варьировать в зависимости от функциональной активности клетки. Наименьшая ширина их может составлять около 20 нм, в расширенном виде они достигают диаметра в несколько мкм.
На внешней поверхности, обращенной к гиалоплазме, находятся рибосомы. Специализируется главным образом на синтезе, транспорте и посттрансляционной модификации белков.
Количество рибосом на ЭР четко связано с его синтетической активностью. Так, на мембранах ЭР в клетке несекретирующей молочной железы связывается до 25% клеточных рибосом, после стимуляции лактации их количество там возрастает до 70%.
Примеры
Клетки печени, где на мембранах шероховатой ЭПС происходит синтез белков плазмы крови (альбуминов, фибриногена, глобулинов, белковых факторов свертывания крови),
плазматические клетки иммунной системы – “фабрики антител”,
экзокринные клетки поджелудочной железы, синтезирующие ферменты для полостного пищеварения.
Роль гранулярного ЭР заключается в процессе сегрегации, обособления синтезированных белков, в их изоляции от основных функционирующих белков клетки.
Котрансляционный синтез растворимых белков
Еще в гиалоплазме происходит связывание иРНК, кодирующей секреторный белок, с рибосомой и начинается синтез белковой цепи. Сначала синтезируется “сигнальная последовательность”, богатая гидрофобными аминокислотами. В нее входит 16-30 аминокислот. На поверхности мембраны ЭР, обращенной к гиалоплазме расположены интегральные рецепторные белки, связывающиеся с большой субъединицей
Такая “заякоренная” рибосома взаимодействует с крупным белковым комплексом. После связывания рибосомы с этим комплексом синтезированный первичный пептид входит в канал диметром около 2 нм. После этого возобновляется синтез полипептида, он удлиняется и его сигнальная последовательность, вместе с растущей цепочкой оказывается внутри полости цистерны ЭР. Таким образом синтезируемый белок проходит сквозь мембрану ЭР во время его синтеза, т.е. котрансляционно, одновременно с его трансляцией.
Внутри полости ЭР с помощью фермента (сигнальная петидаза) сигнальная последовательность отщепляется. После окончания синтеза вся белковая молекула оказывается в полости ЭР и в это время рибосома отделяется от белка и диссоциирует. После этого в интегральном белке канал закрывается.
Во время трансмембранного переноса растущей белковой цепи происходит ее связь с олигосахаридами ( первичное гликозилирование). В полости цистерн ЭР белки претерпевают ряд дополнительных изменений: образуются дисульфидные связи, происходит их правильное сворачивание, происходит сборка третичной структуры белков. Только белки с правильной конформацией в дальнейшем будут переноситься в зону аппарата Гольджи.
Синтез нерастворимых (мембранных) белков
В гранулярном ЭР происходит синтез белков, которые, встраиваясь в мембрану ЭР, становятся интегральными мембранными белками. В цепи синтезирующегося мембранного белка существует одна или несколько аминокислотных стоп-последовательностей,которые препятствуют белковой цепи пересекать мембрану и белок области стоп-сигнала остается связанным с мембраной, нопри этом синтез белка на рибосоме не останавливается. Этоприводит к тому, что весь белок остается встроенным в мембрану. Мембранные белки, также как и растворимые подвергаются различным модификациям.
Первичное гликозилирование
Это ковалентное связывание белковой цепи со сложным олигосахаридом.
В результате этого синтезирующийся белок становится гликопротеидом.
Большинство белков, синтезированных в гранулярном ЭР, относится к гликопротеидам. На белковую молекулу переносится готовый блок олигосахаридов, который связывается с аспарагиновыми остатками белковой молекулы. Этот олигосахаридный комплекс содержит 2 молекулы N-ацетилгликозамина, 9 молекул маннозы и 3 молекулы глюкозы и связан со специальным липидом долихолом на внутренней поверхности мембраны ЭР, смотрящей в просветвакуоли ЭР. По мере транслокации белковой цепи во время ее синтезакаждый аспарагиновый остаток связывается с олигосахаридным комплексом,с помощью фермента, являющегося интегральным белком мембран ЭР.Первичной модификации, гликозилированию, подвергаются как растворимые, так и мембранные белки, синтезирующиеся в ЭР.
Транспорт между ЭР и аппаратом Гольджи
Участки гранулярного ЭР, которые расположены в зоне, приближенной к аппарату Гольджи образуют мембранные выступы, от которых отпочковываются мелкие вакуоли, содержащие синтезированные в ЭР белки. Эта зона называется ЭР-АГ-промежуточный компартмент или везикулярно-тубулярный кластер. Вакуоли, отщепившиеся в этой зоне от ЭР, транзитные элементы, покрыты окаймляющим белковым слоем, аналогичным клатриновому слою эндоцитозных вакуолей
Отделившиеся от ЭР вакуоли становятся окаймленными пузырьками, затем они теряют белковую оболочку и сливаются друг с другом и транспортируются с помощью микротрубочек к цис-зоне аппарата Гольджи, где и сливаются с его мембранами. После деполимеризации окаймляющего слоя на поверхности вакуоли открываются интегральные мембранные белки. Эти белки специфичны для каждого типа вакуолей, направляя их к тому участку, где они должны слиться с другими мембранами. Там они связываются с мембранными белками. В местах взаимодействия этих двух групп белков и происходит слияние мембран.Таким образом происходит транспорт синтезированных белков в зону аппарата Гольджи.
Комплекс Гольджи
(в основной тетрадь)
Ультраструктура
В диктиосоме на расстоянии 20-25 нм друг к другу расположены стопкой уплощенные мембранные мешки, или цистерны, между которыми располагаются тонкие прослойки гиалоплазмы. Каждая отдельная цистерна имеет диаметр около 1 мкм и переменную толщину; в центре ее мембраны могут быть сближены (25 нм), а на периферии иметь расширения, ампулы, ширина которых непостоянна. Количество цистерн в стопке обычно не превышает 5-10.
Кроме плотно расположенных плоских цистерн в зоне АГ наблюдается множество вакуолей. Мелкие вакуоли встречаются главным образом в периферических участках зоны АГ; иногда видно, как они отшнуровываются от ампулярных расширений на краях плоских цистерн.
Морфология
а) По положению и функции, в диктиосомах различают 2 части:
проксимальная (cis-), формирующаяся часть обращена к ЭПС, дистальная (trans-), зрелая - обращена к плазмолемме. Между ними располагается средний или промежуточный участок АГ.
б) К проксимальной части мигрируют пузырьки от гранулярной ЭПС, ‘’обрабатываемые" в диктиосоме полимеры постепенно
Перемещаются от проксимальной части к дистальной и, затем, от дистальной части - отпочковываются секреторные, транспортные везикулы и первичные лизосомы
В дистальном или транс-участке диктиосом к последней цистерне примыкает участок, состоящий из трубчатых элементов и массой мелких вакуолей, часто имеющих фибриллярную опушенность по поверхности со стороны цитоплазмы - это окаймленные пузырьки такого же типа, как и окаймленные пузырьки при пиноцитозе. Здесь происходит разделение и сортировка секретируемых продуктов. Еще дистальнее располагается группа более крупных вакуолей - это уже продукт слияния мелких вакуолей и образования секреторных вакуолей.
(ф-ции)
Модификация белков в аппарате Гольджи
В цис-зону аппарата Гольджи синтезированные в ЭР белки попадают после первичного гликозилирования и редукции нескольких сахаридных остатков. Все белки имеют одинаковые олигосахаридные цепи, состоящие из двух молекул N-ацетилглюкозамина, шести молекул маннозы. В цис-цистернах начинается вторичная модификация олигосахаридных цепей и их сортировка на два класса. В результате олигосахариды на гидролитических ферментах, предназначенных для лизосом (богатые маннозой олгосахариды), фосфорилируются, а олигосахариды других белков, направляемых в секреторные гранулы, или к плазматической мембране, подвергаются сложным превращениям, теряя ряд сахаров и присоединяя галактозу, N-ацетилглюкозамин и сиаловые кислоты.
При этом возникает специальный комплекс олигосахаридов. Такие превращения олигосахаридов осуществляются с помощью ферментов - гликозилтрансфераз, входящих в состав мембран цистерн аппарата Гольджи. Так как каждая зона в диктиосомах имеет свой набор ферментов гликозилирования, то гликопротеиды как бы по эстафете переносятся из одного мембранного отсека в другой и в каждом подвергаются специфическому воздействию ферментов.
Так в цис-участке
происходит фосфорил-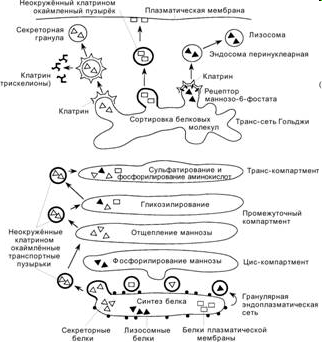
рование манноз в лизосомных ферментах и образуется особая маннозо-6-
группировка, характерная для всех гидролитических ферментов, которые потом попадут в лизосомы.В средней части диктиосом протекает вторичное гликозилирование секреторных белков: дополнительное удаление маннозы и присоединение N- ацетилглюкозамина.
В транс-участке к олигосахаридной цепи присоединяются галактоза и сиаловые кислоты.
Эти данные были получены разными методами. С помощью дифференциального центрифугирования удалось получить раздельные более тяжелые (цис-) компоненты аппарата Гольджи и более легкие (транс-) и определить в них наличие гликозидаз и их продуктов. С другой стороны, используя моноклональные антитела к различным ферментам с помощью электронной микроскопии удалось их локализовать прямо на срезах клеток.
В ряде специализированных клеток в аппарате Гольджи происходит синтез собственно полисахаридов.
В аппарате Гольджи растительных клеток происходит синтез полисахаридов матрикса клеточной стенки (гемицеллюлозы, пектины). Кроме того, диктиосомы растительных клеток участвуют в синтезе и выделении слизей и муцинов, в состав которых входят также полисахариды.
В аппарате Гольджи клеток животных происходит синтез длинных неразветвленных полисахаридных цепей гликозаминогликанов. Один из них, гиалуроновая кислота, входящая в состав внеклеточного матрикса соединительной ткани, содержит несколько тысяч повторяющихся дисахаридных блоков. Многие гликозаминогликаны ковалентно связаны с белками и образуют протеогликаны. Такие полисахаридные цепи модифицируются в аппарате Гольджи и связываются с белками, которые в виде протеогликанов секретируются клетками. В аппарате Гольджи происходит также сульфатирование гликозаминогликанов и некоторых белков.
Слизистые клетки в эпителии секретируют большое количество гликозаминогликанов.
Небольшие пептиды, например некоторые гормоны и нейромедиаторы непосредственно не синтезируются на рибосомах. Они образуются в комплексе Гольджи путём разрезания высокомолекулярного белка-предшественника на короткие пептидные фрагменты.
Сортировка белков в аппарате Гольджи
Через аппарат Гольджи проходит три основных потока синтезированных клеткой нецитозольных белков:
поток гидролитических ферментов в компартмент лизосом,
поток выделяемых белков, которые накапливаются в секреторных вакуолях, и выделяются из клетки только по получении специальных сигналов,
поток постоянно выделяемых секреторных белков.
В цис- и средних зонах диктиосом все эти белки идут вместе разделяясь, для модифиции в зависимости от их олигосахаридных маркеров.
Собственно разделение белков, их сортировка, происходит в транс-участке аппарата Гольджи.
поток гидролитических ферментов в компартмент лизосом
Только белки-предшественники лизосомных гидролаз имеют специфическую олигосахаридную (маннозную группу). В цис-цистернах эти группировки фосфорилируются и дальше переносятся от цистерны к цистерне, через среднюю зону в транс-участок. Мембраны транс-сети аппарата Гольджи содержат трансмембранный белок - рецептор (манноза-6-фосфатный рецептор), который узнает фосфорилированные маннозные группировки олигосахаридной цепи лизосомных ферментов и связывается с ними. На мембранах эти М-6-Ф-рецепторные белки образуют кластеры, группы, которые концентрируются в зонах образования мелких пузырьков, покрытых клатрином.
В транс-сети аппарата Гольджи происходит их отделение, отпочковывание и дальнейший перенос к эндосомам.
Т.е. М-6-Ф-рецепторы, являясь трансмембранными белками, связываясь с лизосомными гидролазами, отсортировывают, от других белков и концентрируют их в окаймленных пузырьках. Оторвавшись от транс-сети эти пузырьки быстро теряют шубу, сливаются с эндосомами, перенося свои лизосомные ферменты, связанные с мембранными рецепторами, в эту вакуоль. Начиная с рН 6 лизосомные ферменты диссоциируют от М-6-Ф-рецепторов, активируются и начинают работать в полости эндолизосомы. Участки же мембран вместе с М-6-Ф-рецепторами возвращаются путем рециклизации мембранных пузырьков обратно в транс-сеть аппарата Гольджи.
поток выделяемых белков, только по получении специальных сигналов
(например нервного или гормонального) проходит такую же процедуру отбора, сортировки на рецепторах транс-цистерн аппарата Гольджи. Эти секреторные белки попадают сначала в мелкие вакуоли тоже одетые клатрином, которые затем сливаются друг с другом. В секреторных вакуолях часто происходит агрегация накопленных белков в виде плотных секреторных гранул. Это приводит к повышению концентрации белка в этих вакуолях примерно в 200 раз, по сравнению с его концентрацией в аппарате Гольджи. Затем эти белки по мере накопления в секреторных вакуолях выбрасываются из клетки путем экзоцитоза, после получения клеткой соответствующего сигнала.
поток вакуолей, связанный с постоянной секрецией
Фибробласты выделяют большое количество гликопротеидов и муцинов, входящих в основное вещество соединительной ткани. Многие клетки постоянно выделяют белки, способствующие связыванию их с субстратами, постоянно идет поток мембранных пузырьков к поверхности клетки, несущие элементы гликокаликса и мембранных гликопротеидов. Этот поток выделяемых клеткой компонентов не подлежит сортировке в рецепторной транс-системе аппарата Гольджи.
Ретроградный перенос везикул.
Путем отщепления от вторичных лизосом вакуолей, которые возвращаются вместе с рецепторными белками в транс-АГ зону. Далее поток вакуолей перемещается от транс-зоны к цис-зоне АГ, а затем от цис-зоны к эндоплазматическому ретикулуму. В этих случаях вакуоли одеты белками COP I-комплекса. Таким путем возвращаются различные ферменты вторичного гликозилирования и рецепторные белки в составе мембран.
В аппарате Гольджи растительных клеток происходит синтез
полисахаридов матрикса клеточной стенки (гемицеллюлозы, пектины).
Диктиосомы растительных клеток участвуют в синтезе и выделении слизей и муцинов.
Комплекс Гольджи не является статической органеллой. При митозе комплекс Гольджи распадается на отдельные кластеры пузырьков и трубочек, распределённых по всей цитоплазме делящейся клетки. К стадии телофазы из этого материала собираются комплексы Гольджи, по одному для каждой дочерней клетки.
Лизосомы
Лизосомы были открыты биохимиком Де Дювом в 1955. При изучении легкой подфракции макросом из гомогенатов печени крысы было найдено, что эта подфракция обладает группой кислых гидролитических ферментов (гидролаз), расщепляющих белки, нуклеиновые кислоты, полисахариды и липиды. Ферменты изолированных лизосом проявляют свою активность только в том случае, если предварительно вызывается повреждение самих лизосом, либо воздействием осмотического шока или детергентов.
(Содержимое)
Наличие некоторых гидролаз можно выявить гистохимическими методами. Одной из характерных гидролаз, является кислая фосфатаза, по наличию которой можно четко определить, является тот или иной мембранный пузырек лизосомой.
(ф-ции)
В некоторых случаях лизосомы могут подходить к плазматической мембране и выбрасывать свое содержимое в наружную среду.
1. у клеток гриба нейроспоры лизосомы, выбрасывая гидролазы из клетки, обеспечивают внеклеточный протеолиз.
2. часть лизосом макрофагов таким же образом обеспечивает внеклеточный гидролиз при воспалительных и резорбционных процессах.
3. При оплодотворении акросома спермия, содержащая гидролитические ферменты гиалуронидазу и протеазы, сливается с плазматической мембраной спермия, изливается на поверхность яйцеклетки. Освободившиеся из вакуоли ферменты расщепляют полисахаридные и белковые оболочки ооцита, давая возможность слиться двум половым клеткам.
(Способы инактивации ферментов)
Лизосомные мембранные белки. В мембрану лизосом встроены специфические белки, в том числе H+,K+‑ATФаза, белки семейства LAMP (LAMP1, LAMP2, CD63/LAMP3). H+,K+‑ATФаза закисляет среду в эндосоме (4,6–5,0), что необходимо для активации лизосомных ферментов. Молекула LAMP2 (Lysosomal-Associated Membrane Protein 2) защищает мембрану лизосомы от самопереваривания и поддерживает в матриксе органеллы кислую среду.
Электронный микроскоп показывает, что фракция лизосом состоит из разнообразных пузырьков размером 0,2-0,4 мкм (для клеток печени), ограниченных одиночной мембраной (толщина ее около 7 нм), с очень разнородным содержанием внутри. Во фракции лизосом встречаются пузырьки с гомогенным, бесструктурным содержимым, встречаются пузырьки, заполненные плотным веществом, содержащим в свою очередь вакуоли, скопления мембран и плотных однородных частиц; часто можно видеть внутри лизосом не только участки мембран, но и фрагменты митохондрий и ЭР. Иными словами, эта фракция по морфологии оказалась крайне неоднородной, несмотря на постоянство присутствия гидролаз.
Типы лизосом
Первичные лизосомы образуются в пластинчатом комплексе в виде одномембранных пузырьков с бесструктурным содержимым. Они предназначены для временного хранения и инактивации гидролаз. Первичные лизосомы способны перемещаться в цитоплазме с помощью микротрубочек, а также сливаться с эндосомами и плазматической мембраной клетки. Диаметр их составляет 100 – 500 нм.
Вторичные лизосомы (фаголизосомы, пишеварительные вакуоли) образуются в результате слияния первичных лизосом с фагоцитарными или пиноцитозными вакуолями. При этом наблюдается активация лизосомальных гидролаз, распад поступивших в клетку путем эндоцитоза веществ и выведение полученных продуктов гидролиза в гиалоплазму для включения в метаболические процессы клетки. Морфологически вторичные лизосомы отличаются от первичных лизосом более крупными размерами и наличием внутри фагоцитируемого материала.
Аутолизосомы морфологически идентичны вторичным лизосомам, отличаясь тем, что содержат внутри другие органоиды клетки (митохондрии, пластиды, рибосомы, включения и т.п.) или их фрагменты. Поэтому они рассматриваются как специальные органоиды, которые обеспечивают аутофагоцитоз. Это явление, часто наблюдается в клетках растений и животных, в частности, при физиологической регенерации внутриклеточных структур и гидролизе запасных питательных веществ.
Остаточные тельца (телолизосомы) представляют собой лизосомы с уплотненным и структурированным содержимым. Они содержат меньше гидролаз и служат местом накопления липидов, пигментов и других продуктов метаболизма.
лизосомные болезни накопления
Болезни накопления характеризуются неспособностью ферментов лизосом (вследствие дефектов их синтеза или структуры) расщеплять метаболиты
Болезнь Дефектный фермент Мишени и типы метаболитов
Болезнь Тэя-Сакса Гексозаминидаза A В нейронах накапливается моносиалоганглиозид
Синдром Хюрлер L-идуронидаза Во многих тканях и органах возрастает
количество гликозаминогликанов
(гепаран сульфат, дерматан сульфат)
Болезнь Помпе (гликогеноз типа 2) Избыточное содержание гликогена
a-1,4-глюкозидаза сердце, скелетных мышцах, печени
(кислая мальтаза) и мозге
Болезнь Ниманна-Пика Сфингомиелиназа Накопление фосфолипидов (главным
образом, сфингомиелина) в
макрофагах печени, селезёнки,
лимфатических узлах и костном мозге
Пероксисомы (микротельца)
Это небольшие вакуоли (0,3-1,5 мкм), одетые одинарной мембраной, отграничивающей гранулярный матрикс, в центре которого располагается сердцевина, или нуклеоид.
В зоне сердцевины часто, особенно в пероксисомах печеночных клеток, видны кристаллоподобные структуры, состоящие из регулярно упакованных фибрилл или трубочек. Изолированные сердцевины пероксисом содержат фермент уратоксидазу .
Пероксисомы обнаружены у простейших (амебы, тетрахимена), у низших грибов (дрожжи), у высших растений в некоторых эмбриональных тканях (эндосперм) и в зеленых частях, способных к фотореспирации, у высших позвоночных животных они обнаруживаются главным образом в печени и почках. В печени крыс на клетку число пероксисом колеблется от 70 до 100.
Пероксисомы часто локализуются вблизи мембран ЭР. У зеленых растений пероксисомы часто находятся в тесном контакте с митохондриями и пластидами.
Впервые пероксисомы были выделены из печени и почек. Во фракциях пероксисом обнаруживается ферменты, связанные с метаболизмом перекиси водорода. Это ферменты (оксидазы, уратоксидаза, оксидаза d-аминокислот) окислительного дезаминирования аминокислот, при работе которых образуется перекись водорода (Н2О2 ) и каталаза, разрушающая ее. В пероксисомах печени каталаза составляет до 40% всех белков и локализована в матриксе. Так как Н2О2 является токсическим веществом для клеток, то каталаза пероксисом может играть важную защитную роль. количество пероксисом в одной клетке может составлять до нескольких сотен. В составе мембраны органеллы находятся специфичные для пероксисом белки — пероксины, а в матриксе — матриксные белки, в т.ч. более 40 ферментов, катализирующие анаболические (биосинтез жёлчных кислот) и катаболические (b-окисление длинных цепей жирных кислот, H2О2‑зависимое дыхание, деградация ксенобиотиков) процессы.
Пероксисомы не содержат никаких нуклеиновых кислот и все белки, из которых они состоят, кодируются ядерными генами, но их относят к саморепродуцирующимся органеллам. В пероксисомах происходит накопление специфических белков, которые синтезируются в цитозоле, и имеют свои сигнальные участки. В мембране пероксисом есть рецепторный белок, который узнает транспортируемые белки.
Формирование пероксисом
Белки для пероксисом транслируются на свободных рибосомах, а затем поступают в органеллу в виде законченных полипептидных цепей. Фосфолипиды, будучи синтезированы в эндоплазматической сети, поступают в пероксисому с участием белков-переносчиков. По мере поступления веществ пероксисомы увеличиваются в размерах. Продолжительность жизни пероксисом — 5–6 суток. Новые органеллы возникают из предсуществующих путём их простого деления.
Глиоксисомы
У животных и некоторых растений (проростки клещевины) играют важную роль при превращении жиров в углеводы. Так, в клетках эндосперма клевещины в глиоксисомах содержатся ферменты глиоксалатного цикла.
Секреторные и транспортные везикулы
Синтез секретов и других веществ происходит, как правило, в мембранных структурах гладкой и шероховатой ПС. Затем они транспортируются в комплекс Гольджи и, после модификации и сортировки, отщепляются от него
Секреторные везикулы выводят вещества из клетки. Бокаловидные клетки кишечника, нейроны.
Транспорные везикулы переносят мембранные белки в плазматическую мембрану
Вакуоли растительных клеток
Клетки как низших, так и высших растительных организмов содержат в цитоплазме вакуоли, несущие ряд важных физиологических нагрузок.
У молодых клеток может быть несколько мелких вакуолей, которые по мере роста и дифференцировки клетки сливаются друг с другом и образуют одну или несколько крупных вакуолей, занимающих до 90% объема всей клетки. Центральные вакуоли отделены от цитоплазмы одинарной мембраной, сходной по толщине с плазмалеммой. Мембрана, ограничивающая центральные вакуоли, носит название тонопласта. Возникают центральные вакуоли из мелких пузырьков, отщепившихся от аппарата Гольджи. Такие первичные вакуоли растут в объеме, сливаются друг с другом и в конце концов образуют одну или несколько крупных вакуолей, оттесняющих цитоплазму с ядром и органоидами к периферии клетки. Полость вакуоли заполнена так называемым клеточным соком, представляющим собой водный раствор, в который входят различные неорганические соли, сахара, органические кислоты и их соли и другие низкомолекулярные соединения, а также некоторые высокомолекулярные вещества (например, белки).
Центральные вакуоли растений выполняют многообразные и важные функции.
-Поддержание тургорного давления клеток. Растворенные в соке вакуолей молекулы определяют его осмотическую концентрацию. Соответствующая молекулярная концентрация сока вакуолей и полупроницаемые свойства как ее мембраны, тонопласта, так и плазмалеммы способствуют тому, что вакуоль функционирует в качестве осмометра и придает клетке необходимую прочность и тургисцентность (напряженность).
- вакуоль представляет собой большую полость, отделенную от метаболирующей гиалоплазмы мембраной, тонопластом, обладающим свойствами полупроницаемости и через котрый может происходить, как и через плазматическую мембрану, активный транспорт различных молекул. В тонопласте обнаружен АТФ-зависимый Н+-насос, направленный внутрь вакуолей, участвующий в транспорте сахаров. Поэтому вакуоли могут использоваться клетками как накопительные резервуары не только для отложения запасных веществ, но и для выброса метаболитов, для экскреции. Так выводятся из клетки все водорастворимые метаболиты. Нерастворимые в воде органические компоненты могут превращаться в растворимые глюкозиды, соединяясь с молекулами сахаров. Перечень экскретируемых в вакуоли метаболитов очень обширен. Это различные алкалоиды (например, никотин, кофеин) и полифенолы. В вакуолях происходит отложение многих глюкозидов, к которым относятся различные пигменты, например антоцианы.
Из неорганических веществ в вакуолярном соке накапливаются фосфаты калия, натрия, кальция, могут накапливаться соли органических кислот (оксалаты, цитраты и др.). Это придает вакуолярному соку отчетливую кислую реакцию (рН от 2 до 5).
Таким образом, тонопласт участвует в процессах экскреции.
- накопление запасных веществ, таких, как сахара и белки. Сахара в вакуолях содержатся в виде растворов, встречаются и резервные полисахариды типа инулина. В вакуолях происходит запасание белков, что характерно для семян. Поступление белков в вакуоли связано со способностью вакуолей ЭР и АГ сливаться с тонопластом. Запасание белков семян злаковых происходит в так называемых алейроновых вакуолях, которые заполняются альбуминами и глобулинами, после чего вакуоли обезвоживаются, превращаясь в твердые алейроновые зерна. При прорастании семян эти зерна обводняются и снова превращаются в вакуоли. В таких новообразованных вакуолях выявляется активность некоторых ферментов, кислой фосфатазы, a-амилазы, глюкозидазы, протеиназы и РНКазы. Следовательно, алейроновые вакуоли отчасти напоминают лизосомы, где происходит переваривание запасных белков при прорастании семян.
Гидролитические ферменты были обнаружены не только в алейроновых вакуолях, но и в мелких и крупных центральных вакуолях. Наблюдалась неоднократно инвагинация, впячивание тонопласта внутрь вакуолей, при этом часть “втянутого” материала оказывается в полости вакуоли и там деградирует. Возможно, так выполняется аутофагическая функция вакуолей, участвующих в гидролизе дефектных клеточных компонентов. Лизосомными свойствами обладают вакуоли дрожжей. Было обнаружено, что стенки вакуолей дрожжей тоже могут образовывать впячивания внутрь, затем они отщепляются от тонопласта и растворяются внутри вакуоли
Двумембранные органеллы
или ПОЛУАВТОНОМНЫЕ
Митохондрии
Митохондрии можно наблюдать в живых клетках, так как они обладают достаточно высокой плотностью. В живых клетках митохондрии могут двигаться, перемещаться, сливаться друг с другом. Размеры митохондрий непостоянны у разных видов, так же как изменчива их форма.
Морфология
Митохондрии или хондриосомы (от греч. mitos - нить, chondrion - зернышко, soma - тельце) представляют собой гранулярные или нитевидные органоиды.
Обычно митохондрии представляют собой мелкие (длиной 0,5-3 мкм, у нитчатых форм 7-60 мкм), у большинства клеток толщина этих структур относительно постоянна (около 0,5 мкм).
Число в клетках разных типов различно.
В клетке печени крысы их около 2500;
В клетках с высокой функциональной активностью (в мотонейронах спинного мозга, в скелетной мышце) число митохондрий превышает десятки тысяч. Биологический полупериод (обновление половины состава) М. в клетках печени составляет 9,6—10,2 суток, в клетках почки — 12,4 суток.
Митохондриальные кристы, отходящие от внутренней мембраны и простирающиеся в сторону матрикса, обычно не полностью перегораживают полость митохондрии, не нарушают непрерывности заполняющего ее матрикса.
риентация крист по отношению к длинной оси митохондрии различна для разных клеток. Так, может быть перпендикулярная ориентация (клетки печени, почек) крист; в некоторых клетках (сердечная мышца) наблюдается продольное расположение крист. Часто кристы могут ветвиться или образовывать пальцевидные отростки, изгибаться и не иметь выраженной ориентации. У простейших, одноклеточных водорослей, в некоторых клетках высших растений и животных выросты внутренней мембраны имеют вид трубок (трубчатые кристы).
Хондриом
Это совокупность всех митохондрий в одной клетке. Во многих клетках хондриом представлен разрозненными многочис-ленными митохондриями, разбросанными равномерно по всей цитоплазме, например, в недифференцированных клетках. В других случаях отдельные митохондрии локализуются группами в местах интенсивной траты АТФ, например, в базальных участках клеток извитых канальцев почки, или в клетках анализаторов сетчатки. В печени выявлены два типа митохондрий: мелкие сферические и крупные разветвленные.
Однако существует и совершенно иной тип хондриома, когда вместо мелких одиночных разрозненных митохондрий в клетке располагается одна гигантская разветвленная митохондрия. Такие митохондрии часто встречаются у одноклеточных зеленых водорослей (Chlorella).
Нитевидные митохондрии длиной около 10 мкм найдены в экзокринных клетках поджелудочной железы. Сперматозоиды содержат цепь длинных митохондрий, состыкованных конец в конец.
Различные типы организации хондриома: а - разрозненные митохондрии, б - нитчатые митохондрии, в - цепочки соединенных митохондрий кардиомиоцитов, г - митохондриальный ретикулум скелетных мышц
Митохондриальная система кардиомиоцитов: а - схема расположения митохондрии и миофибрилл (Ф); ММК - межмитохондриальные контакты; б - флуоресцирующие митохондрии в изолированном кардиомиоците, стрелкой указана одна из митохондрий перед облучением, в - та же клетка после облучения: погасла группа митохондрий, связанных друг с другом с помощью ММК
Митохондрии в скелетной мышечной ткани
Размеры и форма митохондрий весьма разнообразны, но все они имеют две мембраны – гладкую внешнюю и внутреннюю, имеющую кристы, выступающих в сторону матрикса. Мембраны образуют пространственную структуру, в которой расположено множество различных ферментов, индивидуально локализованных на наружной мембране, внутренней мембране, в межмембранном пространстве, в матриксе. Матрикс состоит примерно на 50% из белка; это гомогенная масса с очень тонкой структурой.
Мембраны
Основная структурная особенность митохондрий это наличие у них двух мембран –
наружной
внутренней, которых которая образует
многочисленные впячивания (кристы) в матрикс митохондрии.
Наружная и внутренняя мембраны
Толщина наружной мембраны около 7 нм, она не бывает связана ни с какими другими мембранами цитоплазмы и замкнута сама на себя, так что представляет собой мембранный мешок.
Наружную мембрану от внутренней отделяет межмембранное пространство шириной около 10-20 нм. Внутренняя мембрана (толщиной около 7 нм) ограничивает собственно внутреннее содержимое митохондрии, ее матрикс.
Наиболее многочисленным белком в наружной митохондриальной мембране является порин, который образует большие поры для различных метаболитов. Митохондриальная внутренняя мембрана должна поддерживать электрохимический градиент, следовательно, транспорт метаболитов через эту мембрану четко контролируется, чтобы предупредить утечку ионов. Внутренняя мембрана содержит большое семейство белков переносчиков, которые ответственны за специфическую транслокацию метаболитов.
Состав: Внутреняя мембрана содержит 20-30% липидов, преимущественно кардиолипина, фосфатидилэтаноламина, фосфатидилхолина,инозитфосфатида и фосфатидилсерина;
Дыхание
Дыхание представляет собой большое число последовательных реакций, в ходе которых энергия выделяется маленькими порциями. Весь процесс обычно делят на три этапа: гликолиз, цикл Кребса и окислительное фосфорилирование. Гликолиз происходит в цитоплазме клетки вне митохондрий без участия кислорода, цикл Кребса и окислительное фосфорилирование - внутри митохондрий, окислительное фосфорилирование - при обязательном участии кислорода.
Гликолиз
Глюкоза является обычным субстратом для выработки энергии в клетке, но и другие органические соединения также окисляются,
Жирные кислоты содержат в молекулах четное число атомов углерода и распадаются на двууглеродистые фрагменты, которые при этом должны превратиться в уксусную кислоту. В уксусную же кислоту превращаются и аминокислоты с двумя атомами углерода. Аминокислоты, молекулы которых содержат 3 атома углерода, превращаются в пировиноградную кислоту.
Цикл Кребса (цикл трикарбоновых кислот, цикл лимонной кислоты)
В цикле Кребса участвует ряд кислот:
лимонная (цитрат),
янтарная (сукцинат),
яблочная (малат),
щавелевоянтарная и др.
Поступая на вход цикла Кребса ацетил-КоА проходит через все эти звенья. В конце цикла Кребса получается 3 НАД-Н2 и 1 ФАД-H2. Далее переносчики протонов выходят из цикла Кребса и двигаются к мембране митохондрии. Там начинается третий этап –окислительное фосфорилирование
Окислительное фосфорилирование – один из механизмов образования АТФ
Транспорт электронов в дыхательной цепи сопряжён с образованием протонного градиента, необходимого для синтеза АТФ. Этот процесс называется окислительным фосфорилированием. Окислительное фосфорилировние – это процесс, в котором энергия биологического окисления превращается в химическую энергию АТФ.
В окислительном фосфорилировании участвуют: цитохромы, коэнзим-Q10 (убихинон), АТФ-синтаза, АДФ, остатки фосфорной кислоты, кислород, протоны (НАДН2) и электроны.
Энергию, выделяющуюся при прохождении электрона по дыхательной цепи, цитохромы расходуют на выбрасывание протонов изнутри митохондрии в пространство между ее мембранами. По разные стороны мембраны образуется разность потенциалов между концентрацией протонов, принесённых НАДН2 и электронами с другой стороны, перенесёнными с помощью коэнзима-Q10 на молекулы-цитохромы, размещённые на мембране. Цитохромы прокачивают электроны на противоположенную сторону мембраны, где они соединяются с кислородом.
=>тканевое дыхание представляет собой процесс транспорта электронов и протонов от НАД- или ФАД-зависимых субстратов на кислород, а также протонов, поставляемых матриксом митохондрий. Полное восстановление молекулярного кислорода воздуха в дыхательной цепи сопровождается образованием воды. Дыхательная цепь
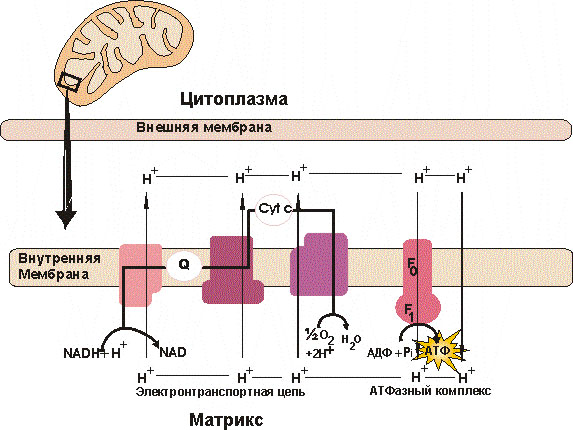
Комплекс I НАДH∙H+-убихинон-оксидодук-таза ответственен за перенос двух электронов от НАДН на убихинон и является местом входа в ЭТЦ восстанов-ленных эквивалентов от НАДН. Разделяет потоки протонов и е
Комплекс II– сукцинат-убихинон-оксидоредуктаза. Перехватывает протоны и е от I и II комплексов ДЦ, и матрикса и пере-Носит их в межмембранное пространство.
Комплекс III– убихинол-цитохром С-Оксидоредуктаза. Осуществляет перенос электронов от убихинона на цитохром с.
Комплекс IV- цитохром С-оксидаза.Переносит протоны из матрикса митохон-дрий в межмембранное пространство.
В межмембранном пространстве создается положительный заряд, а внутри митохондрии - отрицательный. Также на мембране размещены сложные комплексы молекул-ферментов, которые называются АТФ-синтазами. Как только разность потенциалов между электронами и протонами на мембране достигает ~200 мВ, протоны устремляются внутрь матрикса митохондрии - АТФ-синтетаза поворачивается, попутно соединяя АДФ с остатком фосфорной кислоты, в результате чего получается молекула АТФ
(АТФ-азный комплекс).
Единовременно в организме человека продуцируется около 30 грамм АТФ. Именно этот вес иногда называют весом души, т.к. при терминальном состоянии в стадии агонии при отсутствии кровообращения 30 грамм АТФ хватает всего-лишь на 3-5 минут жизни организма
ДНК
У всех эукариот — будь это малярийный плазмодий, разрушающий эритроциты человека, или сам человек, гигантская свободноживущая клетка амеба протей, микроскопическая колония дрожжей или гриб, имеющий многокилометровый мицелий, тысячелетние секвойи — у всех генетическая информация содержится не только в хромосомах клеточного ядра, но и в митохондриях — органеллах клетки, имеющих собственный геном
а) Митохондрии отличаются от прочих органелл двумя интересными особенностями.
Они содержат собственную ДНК - от 1 до 50 небольших одинаковых циклических молекул.
Кроме того, митохондрии содержат собственные рибосомы, которые по размеру несколько меньше цитоплазматических рибосом и видны как мелкие гранулы
б) Данная система автономного синтеза белков обеспечивает
образование примерно 5 % митохондриальных белков.
В 1987 г. А.Уилсон из Калифорнийского университета опубликовал результаты сравнительного анализа ДНК митохондрий, взятых у 147 представителей разных этносов всех человеческих рас, заселяющих пять континентов. По типу, местоположению и количеству индивидуальных мутаций установили, что все митохондриальные ДНК возникли из одной предковой последовательности нуклеотидов
Некоторые субъединичные ферменты дыхательной цепи митохондрий состоят из разных полипептидов, часть которых кодируется ядерным, а часть — митохондриальным геномом.
Цитохром с образуется в гиалоплазме, а из девяти полипептидных цепей в составе АТФ-синтетазы только одна синтезируется в матриксе митохондрий животных.
Набор и количество их генов в мтДНК, для которых полностью определена последовательность нуклеотидов, сильно различаются у разных видов животных, растений, грибов и простейших. Наибольшее количество генов обнаружено в митохондриальном геноме жгутикового простейшего Rectinomonas americana — 97 генов, включая все кодирующие белок гены, найденные в мтДНК других организмов. У большинства высших животных геном митохондрий содержит 37 генов: 13 для белков дыхательной цепи, 22 для тРНК и два для рРНК (для большой субъединицы рибосом 16S рРНК и для малой 12S рРНК).
Геном
Как правило, в каждой митохондрии содержится несколько копий ее генома. Так, в клетках печени человека около 2 тыс. митохондрий, и в каждой из них — по 10 одинаковых геномов. В фибробластах мыши 500 митохондрий, содержащих по два генома, а в клетках дрожжей S.cerevisiae — до 22 митохондрий, имеющих по четыре генома.
Митохондриальный геном растений, как правило, состоит из нескольких молекул разного размера.
Рибосомы
рРНК и рибосомы митохондрий резко отличны от таковых в цитоплазме. Если в цитоплазме выявляются 80S рибосомы, то рибосомы митохондрий растительных клеток принадлежат к 70S рибосомам (состоят из 30S и 50S субъединиц, содержат 16S и 23S РНК, характерные для прокариотических клеток), а в митохондриях клеток животных обнаружены более мелкие рибосомы (около 50S).
Функции митохондрий
Главная функция митохондрий -
завершение окислительного распада питательных веществ и
образование за счёт выделяющейся при этом энергии АТФ - временного аккумулятора энергии в клетке.
1. Состоит из 2 процессов. –
а) Цикл Кребса - распад ацетил-КоА, которым заканчивается разрушение почти всех веществ.
б) Окислительное фосфорилирование - образование АТФ в ходе переноса электронов (и протонов) на кислород.
(Другие процессы, проходящие в митохондриях:) Симбиотическое происхождение митохондрий)
Происхождение
В пользу симбиотической теории говорят многочисленные факты:
— совпадают размеры и формы митохондрий и свободно живущих аэробных бактерий; те и другие содержат кольцевые молекулы ДНК, не связанные с гистонами (в отличие от линейных ядерных ДНК); — по нуклеотидным последовательностям рибосомные и транспортные РНК митохондрий отличаются от ядерных, имея сходство с ДНК и РНК некоторых аэробных грамотрицательных бактерий;
— митохондриальные РНК-полимеразы, хотя и кодируются в ядре клетки, ингибируются рифампицином, как и бактериальные, а эукариотические РНК-полимеразы нечувствительны к этому антибиотику;
— белковый синтез в митохондриях и бактериях подавляется одними и теми же антибиотиками, не влияющими на рибосомы эукариот;
— липидный состав внутренней мембраны митохондрий и бактериальной плазмалеммы сходен, но сильно отличается от такового наружной мембраны митохондрий, гомологичной другим мембранам эукариотических клеток;
— кристы, образуемые внутренней митохондриальной мембраной, являются эволюционными аналогами мезосомных мембран многих прокариот;
— до сих пор сохранились организмы, имитирующие промежуточные формы на пути к образованию митохондрий из бактерий (примитивная амеба Pelomyxa не имеет митохондрий, но всегда содержит эндосимбиотические бактерии).
-Трансляция в митохондриях, как и у бактерий, всегда начинается с особой модифицированной аминокислоты — N-формилметионина. У архей и в цитоплазме эукариот эта аминокислота при синтезе белка не используется. Поэтому наличие N-формилметионина на конце белковой молекулы (или на концах более коротких молекул — формил-пептидов, образующихся при распаде бактериальных белков) является надежным индикатором прокариотического происхождения.
наличие в митохондриальной ДНК (мтДНК) неметилированных участков, содержащих расположенные вплотную друг к другу нуклеотиды Ц и Г . В ядерном геноме млекопитающих такие участки ДНК обычно подвергаются метилированию, но этого не происходит у бактерий и в митохондриях.
Цитоплазматическая (митохондриальная) наследственность
Митохондриальный геном в отдельных случаях участвует в формировании некоторых морфологических и физиологических признаков. К таким признакам относятся характерные для ряда видов высших растений синдром NCS (non-chromosomal stripe, нехромосомно кодируемая пятнистость листьев) и цитоплазматическая мужская стерильность (ЦМС), приводящая к нарушению нормального развития пыльцы. Проявление обоих признаков обусловлено изменениями в структуре мтДНК. При ЦМС наблюдаются перестройки геномов митохондрий в результате рекомбинационных событий, ведущих к делециям, дупликациям, инверсиям или инсерциям определенных нуклеотидных последовательностей или целых генов.
Цитоплазматическая наследственность, в отличие от ядерной, не подчиняется законам Менделя. Это связано с тем, что у высших животных и растений гаметы от разных полов содержат несопоставимые количества митохондрий. Так, в яйцеклетке мыши имеется 90 тыс. митохондрий, а в сперматозоиде — лишь четыре. В оплодотворенной яйцеклетке митохондрии преимущественно только от женской особи, т.е. наследование всех митохондриальных генов материнское, отцовская мтДНК хоть и проникает при оплодотворении в яйцеклетку, однако ее воспроизведение тотчас же блокируется.
Мутации митохондриальной ДНК
С мутациями в митохондриальной ДНК связывают некоторые наследственные болезни и старение человека. Есть данные об участии дефектов мтДНК в канцерогенезе. Следовательно, митохондрии могут быть мишенью химиотерапии рака. Имеются факты о тесном взаимодействии ядерного и митохондриального геномов в развитии ряда патологий человека. Множественные делеции мтДНК обнаружены у больных с тяжелой мышечной слабостью, глухотой, умственной отсталостью, наследующихся по аутосомно-доминантному типу
Заболевания
врожденная атрофия зрительных нервов Лебера (LHON); поражает преимущественно мужчин в возрасте от 16–40 лет; проявляется атрофией сетчатки, двигательными нарушениями ;
KSS — синдром Кернса-Сейра; развивается в молодом возрасте; проявляется пигментной дегенерацией сетчатки,
синдром Пирсона; развивается в детстве; проявляется системными нарушениями окислительного фосфорилирования, поражением центральной нервной системы, костного мозга и поджелудочной железы; как и в случае KSS проявлениям синдрома Пирсона могут предшествовать проявления синдрома де Тони-Дебре-Фанкони;
болезнь Ли; развивается в молодости; проявляется в виде подострой некротизирующей энцефаломиопатии, нейродегенерации;
MERRF — миоклоническая эпилепсия с синдромом «разорванных красных волокон»; характеризуется прогрессирующей миастенией, глухотой;
MELAS — митохондриальная миопатия и энцефалопатия с лактат-ацидозом и инсультоподобными эпизодами; по симптоматике напоминает MERRF; характеризуется генерализованными судорожными припадками, общим истощением и быстрым летальным исходом.
Недавно делеции митохондриального генома были идентифицированы у пациентов с болезнью Паркинсона. Причем в больших количествах: численность дефектных мтДНК у больных паркинсонизмом десятикратно превышала норму.
Помимо паркинсонизма к митохондриальным патологиям сегодня причисляют и болезнь Альцгеймера. Разница лишь в том, что наблюдаемые при болезни Альгеймера дегенерация синаптосомальных митохондрий, снижение активности пируватдегидрогеназы и дефектность митохондриальной дыхательной цепи обусловлены не четко локализованными делециями, а множеством мутаций, «разбросанных» по всему митохондриальному геному.
Иммунная система принимает митохондрии за бактерии.
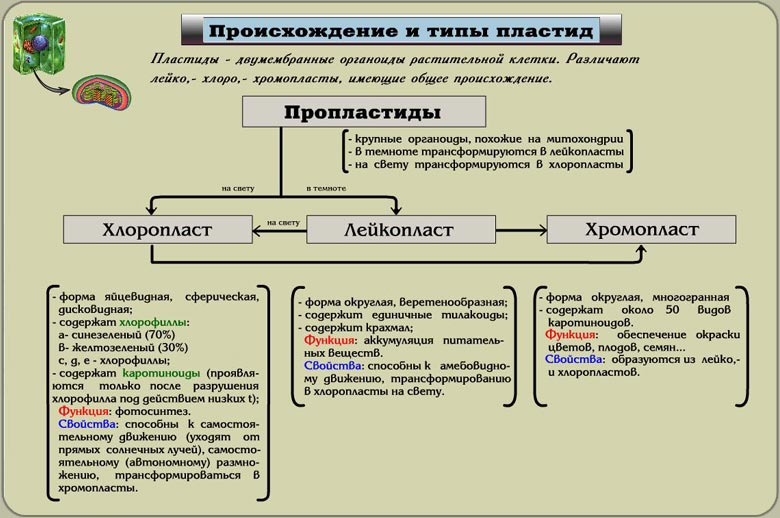
Пластиды - (от греч. plastos - вылепленный; оформленный), органоиды растительных клеток. Содержат пигменты и осуществляют синтез и накопление органических веществ.
Различают зелёные хлоропласты, жёлтые, оранжевые или красные хромопласты и бесцветные лейкопласты. Хлоропласты высших растенийний содержат хлорофиллы и каротиноиды и осуществляют фотосинтез. В лейкопластах из простых органических соединений синтезируются крахмал (амилопласты), масла (элайопласты) и белки (протеинопласты), откладываемые в запас в тканях клубней, корней, корневищ и в эндосперме семян. Хромопласты содержат каротиноиды и придают жёлтую, оранжевую и красную окраску осенним, листьям, лепесткам околоцветника, зрелым плодам (помидоры, рябина, яблоки и др.), корнеплодам (морковь, свёкла и др.).
Хлоропласты
Хлоропласты — тельца линзовидной или округлой формы размером 4—6 мкм (редко до 9 и как исключение до 24 мкм); они содержат около 50% белка, 35% липидов и 7% пигментов
Хлорофилл а –синезеленый (70%)
Хлорофилл в – желто-зеленый (30%)
Хлорофиллы – с,g,е содержат каротиноиды — красно-оранжевый каротин и жёлтый ксантофилл.
ДНК и РНК, рибосомы
Обычно на клетку высших растений приходится в среднем 10—30 хлоропластов. Встречаются клетки с огромным количеством хлоропластов. Например, в гигантских клетках палисадной ткани махорки обнаружено около 1000 хлоропластов.
Внутренняя мембрана хлоропластов отделяет строму пластиды. В строме зрелого хлоропласта высших растений видны два типа внутренних мембран. Это - мембраны, образующие плоские, протяженные ламеллы стромы, и мембраны тилакоидов - плоских дисковидных вакуолей, или мешков.
Ламеллы
Ламеллы стромы (толщиной около 20 мкм) представляют собой плоские полые мешки или же имеют вид сети из разветвленных и связанных друг с другом каналов, располагающихся в одной плоскости. Обычно ламеллы стромы внутри хлоропласта лежат параллельно друг другу и не образуют связей между собой
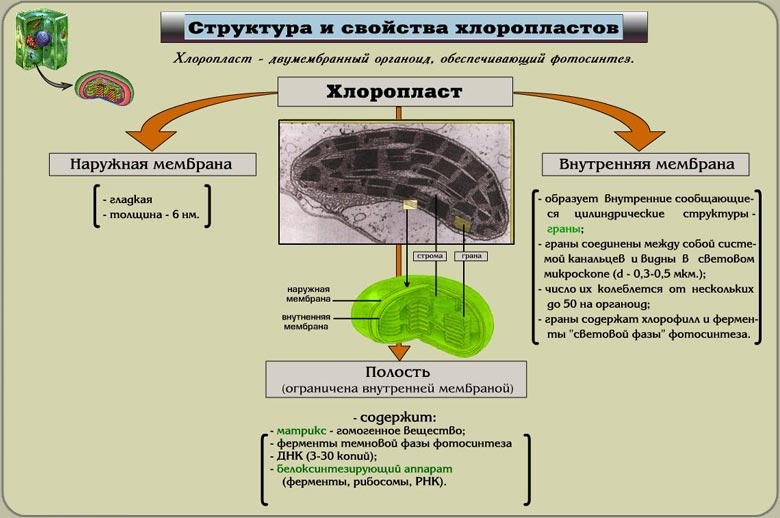
Тилакоиды
Это плоские замкнутые мембранные мешки, имеющие форму диска. Величина межмембранного пространства у них около 20-30 нм. Такие тилакоиды образуют стопки наподобие столбика монет, называемые гранами.
Число тилакоидов на одну грану - от нескольких штук до 50. Размер стопок до 0,5 мкм.
Количество гран до 40-60.
В месте соединения мембран тилакоидов - плотный слой толщиной около 2 нм.
В состав граны также входяти участки ламелл, образуя плотные слои толщиной 2 нм. Ламеллы стромы связывают между собой отдельные граны хлоропласта. Полости камер тилакоидов всегда замкнуты.
На внешней поверхности тилакоидов белковый компонент мембран представлен глобулярными белками-ферментами. В состав мембран входят также хлорофиллы и каротиноиды, образуя липидно-белково-пигментный комплекс, в котором на свету осуществляется фотосинтез. Такое строение во много раз увеличивает их активную синтезирующую поверхность.
Хлорофилл
При помощи хлорофилла зеленые растения поглощают энергию солнечного света и превращают ее в химическую. Поглощение света с определенной длиной волны приводит к изменению в структуре молекулы хлорофилла, при этом она переходит в возбужденное, активированное состояние. Освобождающаяся энергия активированного хлорофилла через ряд промежуточных этапов передается определенным синтетическим процессам
Суммарная реакция фотосинтеза может быть выражена следующим образом:
свет
nCO2 +nH2O → (CH2O)n +nO2
хлорофилл
Световая фаза фотосинтеза
Основные процессы:
Улавливание квантов красного света с помощью ФСI и ФСII.
Процесс фотолиза воды – фоторазложение.
Выделение кислорода в атмосферу.
Основные продукты световой фазы:
О2
НАДФ*Н – восстановленный переносчик
АТФ
Темновая фаза фотосинтеза
Углеродный (пентозный) цикл или цикл Кальвина:
1961 году Мелвин Кальвин объяснил последовательную цепь превращений пентоз, триоз, гексоз, в процессе фиксации СО2. Фиксация одной молекулы СО2 требует затрат трех молекул АТФ и двух молекул НАДФ* Н.
Основные процессы:
Фиксация СО2 из воздуха.
Взаимопревращение углеродных соединений.
Основные продукты темновой фазы:
Глюкоза - С6 Н12О6
АДФ, Ф
НАДФ+
Аминокислоты.
Растения, у которых цикл Кальвина – единственный путь фиксации углерода, называют С 3 – растениями. Растения, которые фиксируют СО2 с образованием четырехуглеродного соединения, называют С4 – растениями. Затем СО2 переносится в цикл Кальвина. С3 – и С4 - путь разделены пространственно – они функционируют в разных клетках. С4 – растения более экономно расходуют СО2 и поглощают его с меньшей потерей воды.С4 – фотосинтез выработался в процессе эволюции у растений жарких, солнечных, засушливых мест, в частности у луговой растительности.У некоторых растений существует еще один путь фиксации СО2 – кислотный метаболизм толстянковых (САМ - растения). Этот путь свойственен кактусам, молочаям и д.р. суккулентам, приспособленным к условиям пустыни. Днем в жаркое время их устьица закрыты и открываются только ночью, когда вода испаряется медленнее. С4 - соединения, запасенные в ночное время, затем в течение дня, когда устьица закрыты, отдают СО2 в цикл Кальвина. Таким образом, у САМ – растений оба пути, осуществляются в одной клетке и разделены по времени.
Лейкопласты
Отсутствует развитая ламеллярная система. Встречаются они в клетках запасающих тканей. Способны к образованию на свету нормальных тилакоидных структур и к приобретению зеленой окраски. В темноте лейкопласты могут накапливать различные запасные вещества, а в строме лейкопластов откладываются зерна крахмала. В некоторых тканях (эндосперм злаков, корневища и клубни) накопление крахмала в лейкопластах приводит к образованию амилопластов
Хромопласт
Имеет желтый свет в результате накопления в нем каротиноидов. Хромопласты образуются из хлоропластов и значительно реже из лейкопластов (например, в корне моркови). Процесс обесцвечивания и изменения хлоропластов легко наблюдать при развитии лепестков или при созревании плодов. При этом в пластидах могут накапливаться окрашенные в желтый цвет капельки (глобулы). При разрушении ламелл хлоропластов выделяются липидные капли, в которых растворяются пигменты (каротиноиды).
Геном пластид
Хлоропласты имеют собственную генетическую систему, обеспечивающую синтез ряда белков. В матриксе хлоропластов обнаруживаются ДНК, РНК и рибосомы Она представлена циклическими молекулами длиной до 40-60 мкм. В одном хлоропласте может быть множество копий ДНК. Так, в хлоропласте кукурузы присутствует 20-40 копий молекул ДНК. ДНК хлоропластов не состоит в комплексе с гистонами. На ДНК хлоропластов синтезируются все виды РНК. ДНК хлоропластов кодирует рРНК, входящую в состав рибосом этих пластид, которые относятся к прокариотическому 70S типу (содержат 16S и 23S рРНК). Рибосомы хлоропластов чувствительны к антибиотику хлорамфениколу подавляющему синтез белка у прокариотических клеток
ДНК может кодировать до 120 генов, среди них: гены 4 рибосомных РНК, 20 рибосомных белков, гены некоторых субъединиц РНК-полимеразы хлоропластов, несколько белков I и II фотосистем, 9 из 12 субъединиц АТФ-синтетазы, части белков комплексов цепи переноса электронов, одной из субъединиц рибулозодифосфат-карбоксилазы (ключевой фермент связывания СО2), 30 молекул тРНК и еще 40 пока неизвестных белков.
Происхождение
Предполагается, что фотосинтезирующие пластиды,как и митохондрии, произошли от свободно живущих бактерий, которые в результате симбиоза были в определенной последовательности включены в состав клеток прокариот - хозяев
Немембранные органеллы
Рибосомы
Количество рибосом - от тысяч до миллионов на клетку
Размер полной рибосомы
про- 20х17х17нм,
эу- 25х20х20нм
Состоят из двух неравных частиц
Не имеют мембраны
Время полужизни рибосом составляет 50-120 часов.
Мембрансвязанные рибосомы
Гранулярная структура ЭПС обусловлена наличием на её поверхности рибосом. Такие рибосомы называются мембраносвязанными; они осуществляют синтез белков, попадающих во внутреннее пространство ЭПС
Рибосомы мембраны ядра
Находятся на поверхности ядра и синтезируют ядерные белки
- Белки ламины
- Белки гистоны
- Негистоновые белки
Мембранонесвязанные
а) или свободные, рибосомы. б) Они синтезируют белки, которые
либо остаются в гиалоплазме, либо переходят в состав тех или иных клеточных структур (митохондрий, цитоплазмы).
в) Содержание таких рибосом особенно возрастает в быстро растущих клетках.
Можно выделить четыре класса рибосом:
- Прокариотические 70S.
- Эукариотические 80S.
- Рибосомы митохондрий (55S - у животных, 75S - у грибов).
- Рибосомы хлоропластов (70S у высших растений).
S - коэффициент седиментации или константа Сведберга. Отражает скорость осаждения молекул или их компонентов при центрифугировании, зависящую от конформации и молекулярного веса.
Митохондриальные рибосомы
Морфология рибосома) Рибосома состоит из двух субъединиц - малой и большой.
1-голова2 – платформа
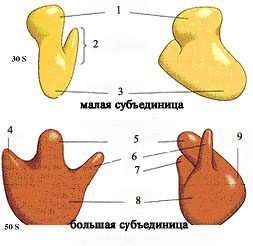 3
- основание4,7- ручка5-центральный
протуберанец6-носик8-передняя часть9-задняя
часть
3
- основание4,7- ручка5-центральный
протуберанец6-носик8-передняя часть9-задняя
часть
(Химический состав)
рибосомные гены эукариот представлены сотнями и тысячами единиц; они принадлежат к фракции умеренно повторяющихся последовательностей ДНК. Общее количество этой фракции ДНК (рДНК) у E. coli составляет около 1% от всей ДНК. У эукариотических организмов этот процент может составлять
0,4 - для человека,
1,3 для дрозофилы,
5,5 для пекарских дрожжей
Количество рибосомных генов на гаплоидный набор хромосомЧеловек - 200Кошка – 1000Мышь – 100Курица – 200Амфиума – 19600Сверчок – 170Кукуруза - 8500
Образование субъединиц
Рибосомные РНК (рРНК) образуются в ядрышках.
Рибосомные РНК обладают сложной вторичной и третичной структурой, образуя сложные петли и шпильки на комплементарных участках, что приводит к самоупаковке.
Чтобы образовались рибосомы необходимо наличие четырех типов рибосомных РНК в эквимолярных отношениях и наличие всех рибосомных белков (все 80 белков разные).
Строение
Каждая из субъединиц - это компактно свёрнутый рибонуклеопротеидный тяж. Такое сворачивание происходит в присутствии белков. р-РНК концентрируется ближе к центру, белки к периферии. Все белки уникальны. Каждая субъединица содержит несколько функциональных центров.
Функциональные центры
Асп-центр специфического узнавания. Малая субъединица. Образует пептидные связи между актами, прочно связывается с рибосомой.
Р-центр - пептидильный, донорный.
Малая субъединица- связывание тРНК присоединенной к растущей полипептидной цепи.
А-центр - аминоацильный, акцепторный. Большая субъединица связывает тРНК несущую следующую добавляемую аминокислоту,
К-центр - каталитический центр. Малая и несколько белков большой субъединицы.
Выходят через ядерные поры в цитоплазму. Дальнейшая сборка субъединиц происходит в цитоплазме в единую рибосому при участии м-РНК и т-РНК (несущей начальную аминокислоту), ГТФ и ионов Mg
Функции
Декодирование матричной РНК. Каждая рибосома полностью прочитывает одну молекулу м-РНК и в соответствии с ее программой синтезирует одну молекулу белка
Формирование пептидных мостиков. Процесс создания химической структуры белка (синтез полипептидной цепи)
Синтез полипептидной цепочки
Инициация
Участвуют отдельные субчастицы. Процесс катализируется IF1,2,3
Малая субчастица рибосомы связывается с мРНК, то есть служит первичным приемником генетической информации для белоксинтезирующего аппарата. После завершения инициации, к ней присоединяется большая субчастица
Элонгация
В процессе элонгации рибосома удерживает мРНК и движется относительно ее в направлении от 5'-конца к 3'-концу. Удержание мРНК на рибосоме - функция малой рибосомной субчастицы, большая субчастица с мРНК не взаимодействует . Участвуют EF
Рабочий цикл рибосомы при элонгации состоит из трех актов:
1) кодонзависимого связывания аминоацил-тРНК (поставляет аминокислоты в рибосому),
2) транспептидации - переноса С-конца растущего пептида на аминоацил-тРНК, т.е. удлинения строящейся белковой цепи на одно звено,
3) транслокации - перемещения матрицы (мРНК) и пептидил-тРНК относительно рибосомы и переход рибосомы в исходное состояние, когда она может воспринять следующую аминоацил-тРНК. Терминация Когда рибосома достигнет специального терминирующего кодона мРНК, синтез полипептида прекращается. При участии факторов терминации (TF) синтезированный полипептид освобождается из рибосомы. После терминации рибосома может повторить весь цикл с другой цепью мРНК или другой кодирующей последователь-ностью той же цепи.
Полисомы
а) С одной цепью мРНК постепенно связывается несколько рибосом.
б) Находясь на равном расстоянии друг от друга, они двигаются по мРНК в одном направлении.
Опорно-двигательный аппарат клетки
Некоторые клетки имеют специализированные структуры, которые позволяют им передвигаться, или перемещать окружающую их жидкость. У многоклеточных животных также есть клетки, способные сокращаться. В основе всех этих двигательных реакций лежат общие молекулярные механизмы. Эти аппараты должны связываться с опорными или каркасными внутриклеточными структурами. Поэтому можно говорить об опорно-двигательной системе клеток
Микротрубочки
Микротрубочки образуют в клетке густую сеть. Сеть начинается от пери-нуклеарной области и радиально распространяется к плазмолемме, следуя за изменениями её формы. Также микротрубочки идут вдоль длинной оси отростков клеток
Строение
а) Стенка микротрубочки состоит из одного слоя глобулярных субъединиц белка тубулина.
б) На поперечном срезе - 13 таких субъединиц, образующих кольцо. Его параметры таковы:
внешний диаметр - dex = 24 нм,
внутренний диаметр - din = 14 нм,
толщина стенки - l стенки = 5 нм.
Длина несколько мкм.
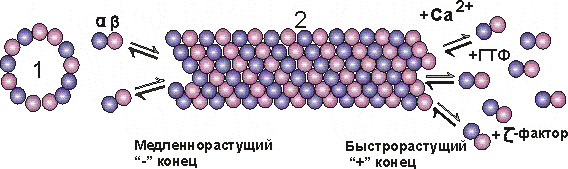
Образование
а)Микротрубочки образуются путём самосборки.
б) Для сборки субъединиц в цитоплазме необходимо присутствие ГТФ, Са 2+, низкомолекулярного τ-белка.
Среднее время полужизни составляет 5 мин. За 15 мин около 80% всей популяции обновляется. Мт веретена деления живут 15-20 сек.
Микротрубочки способны образовывать синглет, дублет и триплет.
микротрубочка A состоит из 13 субъединиц.
Трубочки B и C состоят из меньшего числа протофиламентов, обычно 10
Белки соединяющиеся с микротрубочками. 1. структурные белки и 2. белки транслокаторы.
МАР-microtubulеs-associated proteins
Некоторые MAP соединяют микротрубочки друг с другом, с мембраной или промежуточными филаментами.
Тип I MAP обнаружен в аксонах и дендритах нервных клеток связывает (-)-заряженные участки тубулина.
Тип II MAP также обнаружен в аксонах и дендритах нервных клеток. Присоединяет тубулин.
белки-транслокаторы
(динеины и кинезины), перемещаясь по внешней поверхности микротрубочек, "тащат" за собой органеллы и мелкие пузырьки с транспортируемыми веществами.
Функции
а) В интерфазной клетке создаваемая микротрубочками сеть играет роль цитоскелета, поддерживающего форму клетки.
б) транспорт веществ по длинным отросткам нервных клеток идёт по перитубулярному пространству. Микротрубочки выступают при этом в роли направительных структур
в) В делящихся клетках сеть микротрубочек перестраивается и формирует веретено деления.
Оно связывает хроматиды хромосом с центриолями и способствуют правильному расхождению хроматид к полюсам делящейся клетки.
Некоторые токсины и лекарства, некоторые из которых нарушают митоз влияют на полимеризацию и деполимеризацию тубулина:
таксол - противоопухолевое лекарство, стабилизирует микротрубочки.
колхицин связывает тубулин блокируя полимеризацию. Микротрубочки деполимеризуются при высокой концентрации колхицина.
винбластин - усиливает деполимеризацию.
нокодазол - обеспечивает деполимеризацию микротрубочек.
Производные. Центриоли .
Этот термин был предложен Т. Бовери в 1895 г. (9 х 3) + 0 .
Это значит, что основа центриоли - 9 триплетов микротрубочек, расположенных по окружности, а в центре данного полого цилиндра микротрубочек нет.
Ширина около 0,2 мкм, а длина –
0,3—0,5 мкм
Образование
а) Центриоли располагаются парой - под прямым углом друг к другу. Такая структура называется центросомой.
б) Вокруг центросом - центросфера,
в) Вместе центросома и центросфера называются клеточным центром.
а) В неделящейся клетке - одна пара центриолей.
б) Образование новых центриолей (при подготовке клетки к делению) происходит путём дупликации (удвоения):
каждая центриоль выступает в качестве матрицы,
перпендикулярно которой формируется (путём полимеризации тубулина) новая центриоль.
в) Поэтому одна центриоль является родительской,
а вторая – дочерней
Функции
Индукция полимеризации тубулина при образовании микротрубочек.
В интерфазе в связи с центриолью происходит рост микротрубочек клеточного каркаса.
Центр полимеризации микротрубочек веретена клеточного деления перед митозом .
Центр роста микротрубочек аксонемы ресничек или жгутиков.
Индуцирует полимеризацию тубулинов новой процентриоли, возникающей при ее дупликации
Жгутики
В промежуточной части хвоста - митохондрии (6), расположенные по спирали.
По всей длине хвоста идёт аксонема (5),
Реснички – это покрытые плазмолеммой выпячивания цитоплазмы, по центру которых проходит аксонема.
Имеются в специализированных клетках, в частности, у клеток, выстилающих дыхательные пути (300 тыс. ресничек на одну клетку), в клетке инфузории ~14тыс.
Скорость движения клеток за счет ресничек может достигать ~5мм/c
Реснички
1. Аксонема образована микротрубочками по схеме:
(9 х 2) + 2 .
Т.е.по её окружности расположены девять дуплетов микротрубочек, а ещё пара микротрубочек идёт вдоль оси аксонемы и заключены в центральный футляр.
Схема строения аксонемы реснички.
1A и 1B — A и B микротрубочки периферического дублета,
2 — центральная пара микротрубочек и
центральная капсула,
— центральная пара микротрубочек и
центральная капсула,
3 — динеиновые ручки,
4 — радиальная спица,
5 — нексиновый мостик,
6 — клеточная мембрана.
Функции
Для одноклеточных организмов – движение самой клетки
Для многоклеточных – передвижение веществ по поверхности
Базальное тельце
а) Каждая аксонема прикреплена к базальному телу, находящемуся в поверхностных слоях цитоплазмы.
б) По строению базальное тело похоже на центриоль, т.е. состоит из 9 периферических триплетов. (9х3)+1
в) При этом по две микротрубочки каждого триплета переходят в дуплет аксонемы.
Микрофиламенты
а) Микрофиламенты образуют в клетках густую сеть.В микрофаге – около 100.000 микрофиламентов.
б) основное направление пучков микрофиламентов - вдоль длинной оси клетки и отростков.
Строение
Двойная спираль из глобулярных молекул белка актина.
содержание актина даже в немышечных клетках достигает 10 % от всех белков.
В узлах сети микрофиламентов и в местах их прикрепления к клеточным структурам находятся
- белок a-актинин,
- миозин и тропомиозин.
Образование микрофиламентов
Существуют три формы G–актина a, b и g. В мышечных клетках содержится a–актин, а в немышечных клетках b- и g–актины. Для гладких, сердечных и скелетных мышечных клеток характерны специфичные изоформы a–актина. Важным свойством G–актина является его способность к полимеризации с образованием фибриллярного F–актина.
Сборка микрофиламентов начинается с образования ядра полимеризации. Три молекулы G–актина, несущих АТФ, образуют стабильный комплекс ядро полимеризации, которое служит затравкой для сборки микрофиламента. Полимеризация молекул актина происходит за счёт гидролиза АТФ. При этом в составе микрофиламентов G–актин, несущий продукт гидролиза АТФ (АДФ), остаётся в готовности к быстрой диссоциации (деполимеризации).
Полярность микрофиламентов
полимеризация новых молекул G–актина, несущих АТФ, происходит на (+)–конце, а деполимеризация на (-)–конце микрофиламента.
Известны токсины, связывающиеся с актином и блокирующие его полимеризацию, нарушая тем самым подвижность клеток, фагоцитоз и цитокинез.
Цитохалазины - группа метаболитов плесневых грибов. Они связываются с (+)–концом микрофиламентов, блокируя присоединение новых молекул актина.
Латрункулин морских губок и толитоксин сине-зелёных водорослей (цианобактерий) также останавливают полимеризацию актина.
Фаллоидин (циклический олигопептид бледной поганки Amanita phalloides) в противоположность цитохалазинам, стабилизирует актиновые филаменты, препятствуя как полимеризации, так и деполимеризации
Актин-связывающие белки
с интегральными белками плазмолеммы
α–актинин служит посредником между актином микрофиламентов и мембранными интегринами, обеспечивающими клеточную адгезию. В составе Z-линий и плотных телец ГМК α-актинин фиксирует концы тонких актиновых нитей.
Филамин образует сшивки актиновых филаментов с трансмембранными рецепторами адгезии. В тромбоцитах филамин связывает актиновые нити с интегральным мембранным гликопротеином, что имеет важное значение для прикрепления тромбоцитов к повреждённой стенке кровеносного сосуда.
Белки, сшивающие микрофиламенты
Фимбрин и виллин сшивают F–актин в микроворсинках каёмчатых клеток
Фодрин структурирует терминальную сеть в каёмчатых клетках
Миозин
Миозины — семейство белков, являющихся моторами цитоскелета системы микрофиламентов. Миозины состоят из тяжёлых цепей (H) и лёгких (L) в разном количестве в зависимости от типа миозина. H-цепь имеет 2 участка — «головку» и «хвостик». Головка тяжёлой цепи миозина имеет сайт связывания с актином и сайт связывания АТФ.
Функции
1.Изменение формы клеток (образование псевдоподий) происходит
за счет изменения длины микрофиламентов (в результате дополнительной полимеризации или деполимеризации актина),
за счёт взаимодействия актина с миозином по типу сокращения в мышечных тканях.
Так реализуются следующие формы клеточного движения:
- миграция клеток в эмбриогенезе,
- передвижение макрофагов,
- фаго- и пиноцитоз,
- рост аксонов (у нейронов) и т.д.
2. Примембранный кортикальный скелет. Микрофиламенты образуют скопления по периферии клетки. Актин-связывающие белки прикрепляют микрофиламенты к плазмолемме. Кортикальный скелет поддерживает форму клетки и обеспечивает упругость клеточной мембраны.
3. Изменение консистенции цитозоля, переход золя в гель и обратно (например, для изменения вязкости примембранной цитоплазмы при образовании псевдоподии, в конусе роста аксона).
4. Транспорт. Вместе с микротрубочками микрофиламенты участвуют в направленном транспорте органелл в цитоплазме. Миозин V выполняет роль мотора в быстром аксонном транспорте пузырьков гладкой эндоплазматической сети.
5. Адгезионные фокальные контакты. Микрофиламенты кортикального цитоскелета через α–актинин взаимодействуют с интегринами.
6. Цитокинез (деление цитоплазмы) обеспечивает кольцо из актиновых филаментов вокруг средней части животной клетки.
Микроворсинки
1. а) Микрофиламенты образуют каркас микроворсинок (в тех клетках, где таковые имеются).
б) Это видно на продольных (1) и на поперечных (2) срезах микроворсинок.
2. В основании микроворсинок расположены короткие и толстые нити из белка миозина.
Организация микроворсинки в апикальной части каёмчатой клетки.
Около 30 параллельно идущих микрофиламентов образуют стержень микроворсинки. (+)-концы двух переплетённых нитей F–актина микрофиламентов направлены к вершине микроворсинки. Микрофиламенты сшивают фимбрин и виллин. Микрофиламенты присоединены к внутренней поверхности плазматической мембраны при помощи миозина I. В основании микроворсинок актиновые нити (-)–концами заякорены в терминальную сеть примембранное сплетение микрофиламентов сшитых между собой фодрином
Функционирование
В присутствии АТФ актиновые нити (микрофиламенты) начинают скользить вдоль миозиновых и втягиваться в клетку.
Это способствует перемещению в клетку содержимого микроворсинок
при всасывании веществ в просвете кишечника или
и реабсорбции веществ из канальцев почек
Промежуточные филаменты
Третий компонент цитоскелета - промежуточные филаменты .
- диаметр 8–12 нм - имеют тканеспецифическую природу.
Встречаются во всех типах клеток, особенно в тех, которые подвержены механическим воздействиям. В клетках растений не обнаружены.
2. Промежуточные филаменты часто располагаются параллельно поверхности клеточного ядра
Состоят из мономеров фибриллярных белков, принадлежащих к одному семейству генов. Экспрессия белков промежуточных нитей специфична для определённых клеточных типов.
В состав ПФ входит большая группа изобелков,
1. В эпителии они образованы белком кератином,
2. в клетках соединительной ткани - виментином,
3. в гладких мышечных клетках - десмином,
4. в астроцитах – глиальный белок
5. в нервных клетках они называются нейрофиламентами и тоже образованы особым белком.
6.в ядре – белки ядерной ламины
Белки промежуточных филаментов различных клеток
Белок |
Локализация |
Некоторые специальные функции |
Цитокератин (29 изоформ) |
Эпителиальные клетки |
Создают тянущее усилие, взаимодействуют с десмосомами и полудесмосомами. Маркёры опухолей эпителиального происхождения (плоскоклеточная карцинома, аденокарцинома) |
Десмин |
Клетки мышечных тканей |
Структурируют сократительные органеллы миофибриллы. Маркёры опухолей мышечного происхождения (рабдомиосаркома) |
Виментин |
Клетки мезенхимного генеза: фибробласты, ГМК, эндотелиальные, хондробласты, остеобласты, макрофаги |
Формируют сетевидную структуру вокруг ядра клетки. Маркёры опухолей соединительной ткани (фибросаркома, липосаркома, ангиосаркома, хондросаркома, остеосаркома) |
Глиальный фибриллярный кислый белок |
Астроциты, олигодендроциты, шванновские клетки |
Структурная поддержка цитоплазмы. Маркёры опухолей, возникающих из глиальных клеток (астроцитомы) |
Белки нейрофиламентного триплета |
Нейроны |
Поддерживают структуру и форму отростков нейрона (аксонов и дендритов) |
Ламины A, B и C |
Ядерная пластинка |
Организуют ядерную пластинку и лежащий около неё хроматин |
Цитоплазматические ПФ являются самыми ста-бильными и долгоживу-щими элементами цито-скелета. Они являются истинной опорой клеткам, испытывающим значитель-ные механические нагруз-ки. В клетках эпидермиса они образуют пучки и создают жесткую внутри-клеточную сеть
Включения
а) Включения - необязательные компоненты цитоплазмы; они возникают и исчезают в зависимости от состояния клетки.
Включения могут быть различной природы, но в животной клетке никогда не накапливается белок
ЯДРО
термин «ядро» впервые был применен Брауном в 1833 г. Ядро - важнейшая составляющая клетки, содержащая наследственный материал - ДНК.
Поэтому в соматических клетках оно выполняет 2 ключевые функции:
1. сохраняет наследственный материал для передачи дочерним клеткам (образующимся при делении исходной);
2. обеспечивает использование информации ДНК в самой клетке - в том объёме, в каком это необходимо данной клетке при данных условиях.
Морфология
ДНК каждой клетки содержит следующую информацию:
- о первичной структуре (последовательности аминокислот) всех белков всех клеток организма (исключение - некоторые белки митохондрий, кодируемые митохондриальной ДНК),
- о первичной структуре (последовательности нуклеотидов) примерно 60 видов транспортных РНК и 5 видов рибосомных РНК,
- о программе использования данной информации в разных клетках в разные моменты онтогенеза.
Процессы происходящие в ядре
ключевая функция ядра (использование информации ДНК для обеспечения клеточной жизнедеятельности) реализуется за счёт того, что в нём проходят
- транскрипция определённых участков ДНК (синтез пре-мРНК),
- созревание мРНК,
- синтез и созревание тРНК и рРНК.
Кроме того, в ядре
-формируются субъединицы рибосом (из рРНК и поступающих из цитоплазмы рибосомальных белков).
перед делением клетки (кроме второго деления мейоза) в ядре происходит репликация (удвоение) ДНК, причём в дочерних молекулах ДНК одна из цепей является старой, а вторая - новой (синтезированной на первой по принципу комплементарности).
Структура ядра
3 основных элемента:
- ядерная оболочка,
- глыбки хроматина,
- округлые ядрышки.
б) Другие компоненты ядра -
- ядерный матрикс и
- ядерный сок -формируют ту среду, в которой находятся хроматин и ядрышко.
Ядерная мембрана
Роль ядерной оболочки в ядерно-цитоплазматическом обмене
Регулятор в ядерно-цитоплазматическом обмене. В этом процессе комплекс ядерной поры выступает как супрамолекулярный комплекс, выполняющий роль транслокатора, и сортировщика.
1. Ядерные поры пропускают частицы определенного размера пассивно, по градиенту концентрации. Ионы, нуклеотиды, АТФ, гормоны - свободно поступают в ядра.
2. Через ядерную оболочку беспрепятственно в обе стороны происходит пассивный транспорт высоко молекулярных соединений, имеющих массу не более 5 х 103 дальтон.
3. С другой стороны ядерные поры осуществляют избирательный транспорт. Многие белки поступают как в ядро, так и выходят из него против градиента концентраций (гистоны).
4. многие ядерные белки проходят через ядерные поры с помощью специальных механизмов, включающих узнавание и связывание крупных ядерных белков, а затем только их транслокацию, перенос через поры.
Особенности строения
1. Наличие двух мембран.
Ядерная оболочка образована не одной, а двумя мембранами - внешней (1) и внутренней (2), - которые разделены перинуклеарным пространством (3).
В результате ядерная оболочка образует полый двуслойный мешок.
2.Ядерный поровый комплекс (ЯПК или NPC - nuclear pore complex)
структура с молекулярным весом более 125х106 Да, состоящую из более 1000 белков. Белки ЯПК носят название нуклеопоринов 50-100 видов Диаметр около 100 нм.
Комплекс включает: канал;
- белковые гранулы, расположенные по периферии,
- центральную белковую гранулу, которая связана фибриллами с периферическими гранулами.
Количество пор в ядерной оболочке тем больше, чем интенсивней идут в клетке синтетические процессы.
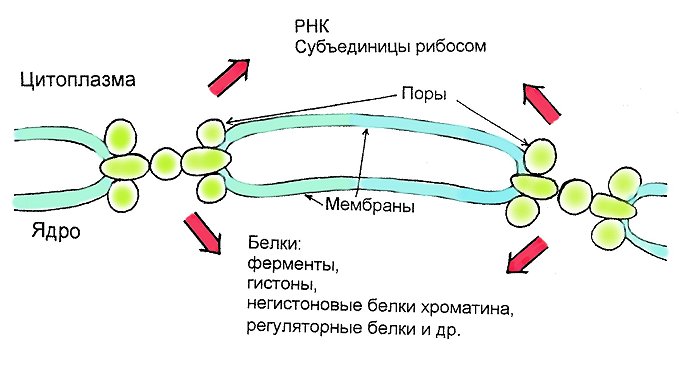
Ядерный матрикс
Внутренняя мембрана связана с фиброзным слоем, ядерной ламиной (Lamina nucleum limitans), которая, заякоривает хроматин на ядерной оболочке.
Фиброзный слой ламины все время перестраивается, особенно в связи с ростом поверхности ядра, во время клеточного цикла.
Ламины A, C и B - промежуточные филаменты, толщиной около 10 нм. Со стороны кариоплазмы под внутренней ядерной мембраной образуют ортогональные структуры, чередующиеся с рыхло расположенной сетью этих же фибрилл.
Ядерный сок (кариоплазма,кариолимфа, нуклеоплазма)
основная внутренняя среда ядра, заполняющее пространство между хроматином, ядрышком и другими структурами. Вязкость ядерного сока примерно такая же, как вязкость основного вещества цитоплазмы.
Кариоплазма имеет вид гомогенной или мелкозернистой массы с низкой электронной плотностью. Вода - 80-85%
Содержит различные ферменты, нуклеотиды, аминокислоты и другие вещества, необходимые для обеспечения синтеза нуклеиновых кислот и субъединиц рибосом.
Из простых белков (протеинов) в кариоплазме содержатся гистоны, протамины, альбумины и глобулины, а из протеидов - липопротеиды, глюкопротеиды и нуклеопротеиды. Большая часть белков относится к глобулярным, меньшая - к фибриллярным структурам
Функции
связаны коллоидные свойства ядра,
роста
синтез ДНК, различных РНК и белка,
передача раздражения
ДНК
На основании биохимических исследований и расчётов установлены следующие характеристики ядерной ДНК.
а) В ядре любой соматической клетке содержится 46 молекул ДНК разной длины - по одной молекуле в каждой из 46 хромосом
б) Средняя длина одной молекулы - 4 см (120.000.000 нуклеотидных пар)всех вместе (в 1 ядре) - около 2 м.
в) Общая масса всей этой ДНК (в 1 ядре) - 5,7 пг (5,7x10-12г),во всех клетках организма человека - около 200 г.
Хроматин
а) Хроматин занимает основную часть объёма ядра.
б) Он представлен (1) тёмными (электроноплотными) глыбками - т.н. гетерохроматином (2) светлыми (электронопрозрачными) областями - эухроматином.
в) Причём, глыбки гетерохроматина находятся, главным образом, на периферии ядра и прилежат к ядерной оболочке (3).
Состав хроматина
а) Весь хроматин в целом - это совокупность 46 хромосом (у человека).
б) Каждая из них представляет собой нуклеопротеидный комплекс - двухцепочечную молекулу ДНК, которая определённым образом связана с ядерными белками.
в) Содержание белков в хроматине по массе в 1,3-1,7 раза больше, чем ДНК.
г) Кроме того, в хроматине обнаруживается и-РНК (являющаяся продуктом транскрипции).
Эухроматин - это функционально активные (участвующие в транскрипции) части хромосом, которые находятся в деконденсированном (диффузном) состоянии.
Гетерохроматин - функционально неактивные отделы, которые конденсированы, образуя глыбки.
При изменении состояния клетки или в процессе дифференцировки возможен переход части гетерохроматина в эухроматин и обратно (факультативный гетерохроматин(Х-хромосома в организме человека)).
Конститутивный гетерохроматин генетически не активен, он не транскрибируется, реплицируется он позже всего остального хроматина, в его состав входит ДНК, с высокоповторяющимися последовательностями нуклеотидов; он локализован в центромерных, теломерных зонах митотических хромосом.
Доля конститутивного хроматина может быть неодинакова. У млекопитающих на него приходится 10-15% всего генома, у амфибий –до 60%. Функциональное значение структуризация интерфазного ядра, с некоторыми регуляторными функциями.
1. чем больше в ядре доля гетерохро-матина, тем ниже функциональная активность ядра
2. а) в ядре нервной клетки (I) гетерохроматина очень мало - ядро и клетка в целом функционально очень активны.
б) в лимфоците (II) наблюдается преобладание гетерохроматина. Это вполне коррелирует с очень малым объёмом цитоплазмы, которая к тому же бедна органеллами. Данная клетка циркулирует в крови, и процессы синтеза РНК и белков идут в ней с небольшой скоростью.
В среднем в хроматине около 40% приходится на ДНК и около 60 % на белки, среди которых специфические ядерные белки-гистоны, составляют от 40 до 80% от всех белков, входящих в состав выделенного хроматина. РНК, углеводы, липиды, гликопротеиды.
Гистоны
Эукариотические клетки содержат 5-7 типов молекул гистонов
Н1,H2A и H2B, Н3 и Н4
гистоны H3 и H4 являются наиболее эволюционно консервативными
Гистон H1 - класс белков, состоящих из нескольких близкородственных белков с перекрывающимися последовательностями аминокислот. Характерны межвидовые и межтканевые вариации
Негистоновые белки
составляют около 20% от всех белков хроматина. Это сборная группа белков, отличающихся друг от друга как по общим свойствам, так и по функциональной значимости.
Во фракцию негистоновых белков может входить около 450 индивидуальных белков с различной молекулярной массой (5-200 кД).
Среди негистоновых белков обнаруживается целый ряд регуляторных белков как стимулирующих инициацию транскрипции, так и ингибирующих ее, обнаружены белки специфически связывающиеся с определенными последовательностями на ДНК. К негистоновым белкам относят также ферменты, участвующие в метаболизме нуклеиновых кислот (ДНК-полимеразы, ДНК-топоизомеразы, метилазы ДНК и РНК, РНК-полимеразы, РНКазы и ДНКазы и т.д.), белков хроматина (протеинкиназы, метилазы, ацетилазы, протеазы и др.) и многие другие.
Уровни упаковки ДНК в хромосомах
Общая длина всей ДНК в ядрах клеток человека примерно равна его среднему росту – 175 см, а диаметр ядра средней клетки составляет около 10 микрон. Это значит, что ДНК должна быть укорочена почти в миллион раз. Различают уровни упаковки (компактизации) ДНК в хромосомах:
уровень нуклеосомы; ДНК укорачивается примерно в 7 раз
уровень нуклеомера; в 6 раз
уровень хромонемы; в сотни и даже многие тысячи раз
уровень хроматиды; в 3 раза
уровень хромосомы. в 90 раз
Первый уровень
Спираль ДНК
Второй уровень – Нуклеосома
Благодаря гистонам, хроматин имеет нуклеосомную организацию.
при электронной микроскопии препаратов хроматина можно видеть, что нити хроматина представляли собой “бусы на нитке”:
а) Основа каждой нуклеосомы
глобула (около 10 нм) из 8 молекул гистонов (октамер).
б) Двуцепочечная молекула ДНК последовательно "намотана" на огромное количество таких глобул,
делая вокруг каждой из них 1,75 оборота
Каждая глобула, связана друг с другом отрезками ДНК длиной около 20 нм
Октамер гистонов образует белковую основу-сердцевину (кор), по поверхности которой располагается ДНК величиной в 146 п.н., образующая 1,75 оборота; остальные 54 п.н. ДНК образуют участок, несвязанный с белками сердцевины - линкер, который, соединяя две соседние нуклеосомы, переходит в ДНК следующей нуклеосомы. Следовательно, полная нуклеосома содержит около 200 п.н. ДНК, октамер сердцевинных (коровых) гистонов и одну молекулу гистона H1. Молекулярная масса полной нуклеосомы - 262000 Да. На весь гаплоидный геном человека (3 х 109 пар оснований) приходится 1,5 х 107 нуклеосом.
Третий уровень – соленоидный или нуклеомерный (супербидный)
Нить плотно упакованных нуклеосом диаметром 10 нм образует спиральные витки с шагом спирали около 10 нм. На один виток приходится 6 нуклеосом. В результате такой упаковки возникает фибрилла спирального типа с центральной полостью, которая иногда на негативно окрашенных препаратах бывает видна как узкий «канал» в центре фибриллы. Гистон H1 обеспечивает взаимодействие между соседними нуклеосомами, не только сближая и связывая их друг с другом, но и обеспечивая кооперативную связь нуклеосом так, что образуется довольно плотная спираль из 30 нм фибриллы
Соленоидный уровень укладки хроматина обеспечивает 40 кратное уплотнение ДНК, Второй уровень компактизации ДНК играет роль фактора, инактивирующего гены.
Четвертый уровень- Петлевые домены ДНК
Образование третьего уровня связано с негистоновыми белками. специфические белки связываются с особыми участками ДНК, которые в местах связывания образуют большие петли или домены. Этот уровень компактизации ДНК связаны не с ее дополнительной спирализацией, а с образованием поперечной петлистой структуры, идущей вдоль интерфазной или митотической хромосомы.
Количество петель в такой розетке может составлять 15-80, а общая величина ДНК может достигать 200 т.п.н., с суммарной длиной ДНК до 50 мкм.
Розетки фибрилл хроматина – хромомеры - приводят уже к 600-кратной компактизации ДНК
300 нм-фибриллы дополнительно сворачиваются, образуя хроматиды диаметром примерно 600-700 нм

Пятый и шестой уровень – хроматидный и хромосомный
Диаметр равен 1400 нм.
Метафазная хромосома уже удвоена.
Она состоит из двух хроматид. Каждая из них содержит одну молекулу ДНК.
Сюда входят белки ядерной ламины,серия белковых нитей, сопряженных с
ядерной оболочкой и пронизывающих
все ядро.
Схема строения хромосомы в поздней
профазе -метафазе митоза. 1-хроматида; 2-центромера; 3- короткое плечо; 4-длинное плечо
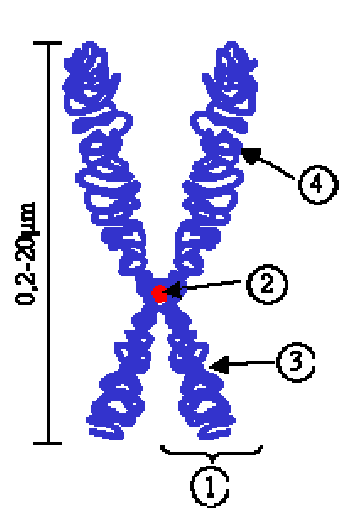
Ядрышко
а) Самая плотная структура ядра - это ядрышко, обычно имеющее округлую форму.
б) В ядре может содержаться несколько ядрышек
Природа
Ядрышко - это производная от хроматина структура.
а) Оно формируется в связи с определёнными участками хромосом - т.н. ядрышковыми организаторами. б) Каждый такой организатор содержит несколько сотен копий генов рибосомной РНК.
а) На этих генах активно происходит синтез предшественников рРНК. б) Последние тут же (в ядре) подвергаются созреванию и, связываясь с рибосомальными белками, образуют субъединицы рибосом (которые выходят из ядра в цитоплазму).
Размер ядрышка отражает степень его функциональной активности, которая широко варьирует в различных клетках и может изменяться в индивидуальной клетке.
В ядрышке происходят транскрипция рибосомных генов, процессинг предшественников рРНК и сборка прерибосомных частиц из рибосомных белков и рРНК. Механизмы формирования ядрышка не ясны. В соответствии с одной из гипотез, ядрышко рассматривают как нуклеопротеиновый комплекс, спонтанно появляющийся в результате объединения регуляторных белково-нуклеиновых комплексов, возникающих на повторяющихся последовательностях рДНК во время их транскрипции. Действительно, гены рРНК человека организованы в виде 250 тандемно повторяющихся последовательностей длиной в 44 т.п.о. каждая, которые вместе с ассоциированными с ними белками формируют сердцевину ядрышка. Оно заполняется другими компонентами во время процессинга рРНК и сборки рибосомных субчастиц.
Ультраструктура ядрышка
Морфологически в ядрышке различают три основные зоны: фибриллярный центр , окруженный плотной фибриллярной и гранулярной областями
1) слабоокрашивающийся компонент, содержащий ДНК из области ядрышкового организатора хромосомы.
Здесь локализованы гены рРНК , РНК-полимераза I , транскрипционный фактор UBF.
Является местом сборки регуляторных нуклеопротеиновых комплексов, необходимых для транскрипции генов рРНК.
2) плотный фибриллярый компонент, состоящий из множества тонких (5 нм) рибонуклепротеиновых фибрилл, представляющих собой РНК-транскрипты
Окружает центр ядрышка, представлен растущими цепями предшественников рРНК и ассоциированными с ними белками, участвующими в процессинге.
3) гранулярный компонент, в состав которого входят частицы диаметром 15 нм, представляющие наиболее зрелые предшественники рибосомных частиц.
В гранулярной области ядрышка обнаруживают зрелые 28S и 18S рРНК, частично процессированные РНК, а также продукты сборки рибосомных субчастиц.
Предшественники рибосом представлены частицами диаметром 15-20 нм. Перенос прерибосомных субчастиц к цитоплазме обеспечивают специфические белки, которые перемещаются от ядрышка к оболочке ядра.
Клеточный цикл
Клеточный цикл – это совокупность всех морфологических и физиолого-биохимических событий, происходящих в клетке от деления до деления (включая деление) или от деления до гибели
В клеточном цикле и выделяют два основных этапа:
1) интерфаза;
2) период клеточного деления.
Длительность клеточного цикла в размножающихся клетках составляет 10 – 50 часов и зависит от типа клеток, их возраста, уровня гормонального баланса, времени суток, физико-химических и ряда других параметров. Клетки быстро делящихся тканей называют активно пролиферирующими.
Пролиферация (от лат. пролес – отпрыск, потомство и феро – несу) – это увеличение числа клеток путем митоза, ведущее к росту ткани и органа.
1. Клеточный цикл – состоит из интерфазы и митоза
Интерфаза состоит из 3 периодов:
G1 - пресинтетический (постмитотический).
S - синтетический,
G2 - постсинтетический (или премитотический),
+ M - митоз,
Контрольные точки клеточного цикла
1. Точка выхода из G1-фазы, называемая
Старт - у млекопитающих и точкой рестрикции у дрожжей. После перехода через точку рестрикции R в конце G1 наступление S становится необратимым, т.е. запускаются процессы ведущие к следующему делению клетки.
2. Точка S – проверка точности репликации.
3. Точка G2/M-перехода – проверка завершения репликации.
4. Переход от метафазы к анафазе митоза
Интерфаза
Исторически название интерфаза (т. е. промежуток между фазами), означало период, в ходе которого в клетке не видно никаких преобразований между предшествующей телофазой и наступающей профазой. Биохимические исследования первой половины ХХ века продемонстрировали, что все основные метаболические процессы, синтез всех биополимеров происходит, тем не менее, в интерфазе, причем более или менее равномерно
Интерфаза объединяет общим термином стадии G1, S и G2
G1-период
а) Период G1- это интервал времени
- от окончания митоза - до начала синтеза ДНК (и ядерных белков) в дочерней клетке.
б) В этот период происходят
- Синтез рибосом, восстановление содержания цитоплазматических белков
- Удвоение митохондрий, синтез АТФ
- Восстановление мембранных структур и,
- как следствие, рост объема цитоплазмы клетки (до размера материнской).
в) Содержание ДНК в клетке - 2n.
У лимфоцитов тимуса составляет менее 3 часов, в кишечном эпителии – 10 часов, в печени -48 часов, в эпидермисе кожи – 64 часа.
S-период
а) В S-период происходят
в ядре - удвоение (репликация) ДНК и увеличение вдвое количества хромосомных белков а возле ядра - дупликация центриолей.
б) В клетках, находящихся на этой стадии, обнаруживается разное количество ДНК - от 2n до 4n, по мере хода синтеза (репликации) ДНК.
для лимфоцитов тимуса 6 часов, в кишечном эпителии – 8 часов, в печени – 16 часов, в эпидермисе кожи – 8 часов.
G2-период
а) Следующий период - G2 - обычно не очень продолжителен и включает синтез ряда других веществ, необходимых для прохождения митоза;среди этих веществ - белок микротрубочек тубулин, используемый для формирования веретена деления.
б) Содержание ДНК в этот период - 4n.
Продолжительность G2-периода у разных клеток меняется мало, составляя в большинстве случаев 2-4 часа.
МИТОЗ (непрямое деление)
Впервые фазы митоза были описаны русским ботаником И.Д. Чистяковым в 1874 году. Но систематические детальные исследования были впервые выполнены на растениях Э. Страсбургером (1876-1879), а на животных – В. Флеммингом (1882). Длительность митоза 1–2 часа.
Состоит из четырех фазы: профаза, метафаза, анафаза и телофаза.
Значение митоза
Генетическая стабильность.В результате митоза получаются два ядра, содержащие каждое столько же хромосом (диплоидный набор), сколько их было в родительском ядре. Эти хромосомы происходят от родительских хромосом путем точной репликации ДНК, поэтому гены их содержат совершенно одинаковую наследственную информацию. Дочерние клетки генетически идентичны родительской клетке, так что никаких изменений в генетическую информацию митоз внести не может. Поэтому клеточные популяции(клоны), происходящие от родительских клеток, обладают генетической стабильностью
Профаза
Профаза Обычно самая продолжительная фаза клеточного деления. Хроматиды укорачиваются ( до 4% своей первоначальной длинны ) и утолщаются в результате их спирализации и конденсации. При окрашивании хроматиды четко видны, но центромеры не выявляются.
Прометафаза (14-60 мин)
У животных клетках и у низших растений центриоли расходятся к противоположным полюсам клетки. От каждой центриоли в виде лучей расходятся короткие микротрубочки, образуюшие в совокупности звезду. Ядрышки исчезают, так как их РНК частично переходит в определенные пары хроматид. К концу профазы ядерная мембрана распадается и образуется веретено деления.
Динамика ядерной мембраны
При этом в профазе по мере конденсации хромосом ядерная оболочка теряет с ними связь, а затем в ней появляются разрывы. Она приобретает вид плоских мембранных вакуолей, цистерн. В это время ядерные поры еще видны. Позднее они исчезают. Во время митоза 120 мДа комплекс ядерной поры разбирается на субкомплексы примерно по 1 мДа.
Метафаза (5-30 мин)
На стадии метафазы в клетке отсутствует ядерная оболочка. Хромосомы
-становятся максимально конденсированными,
-выстраиваются в экваториальной плоскости клетки, образуя метафазную пластинку ,
- в конце метафазы разделяются на 2 хроматиды, которые остаются связанными только в области центромерных перетяжек
В веретено входят микротрубочки 3-х видов:
-кинетохорные: связывают каждую хроматиду (в области её кинетохоры)
-полярные: идут от одной из центросом центру веретена, где перекрываются с микротрубочками от другого полюса;
- полюсные (микротрубочки сияния): направлены к поверхности клетки.
Ядерная оболочка превращается в скопление мелких мембранных пузырьков, окружающих зону бывшего интерфазного ядра. Такие пузырьки морфологически нельзя отличить от других мелких вакуолей в цитоплазме. В метафазе мембранные элементы цитоплазмы оттесняются к периферическим зонам клеток микротрубочками веретена деления.
Анафаза (5-20 мин)
а) Хроматиды, сохраняя максимальную степень конденсации, теряют связь друг с другом и начинают расходиться к полюсам клетки.
б) При этом они ориентированы
-центромерными участками - к соответствующему полюсу,
- теломерными (концами) - к экватору клетки.
в) Одна из хроматид отходит к одному, а другая - к противоположному полюсу.
Поэтому дочерние клетки получают полные и равные наборы хромосом.
В конце анафазы, когда прекращается движение хромосом к противоположным полюсам клетки, мембранные пузырьки цитоплазмы, и в первую очередь мембраны гранулярного эндоплазматического начинают контактировать с поверхностью хромосом. Участки растущих плоских мембранных мешков сливаются, замыкая и отгораживая содержимое нового интерфазного ядра. Ядерные поры появляются на самых ранних этапах реконструкции ядерной оболочки.
Телофаза
1. Набор расходящихся хромосом, приблизившись к полюсам останавливается.
2. Вокруг него начинает формироваться ядерная оболочка.
3. Хромосомы постепенно деконденсируются.
4. В ядрах начинают формироваться ядрышки.
а) Затем между ядрами происходит
- разделение тела клетки (цитотомия, цитокинез)
б) по экватору клетки формируется актомиозиновое кольцо, которое постепенно сжимается, стягивая за собой плазмолемму -
- образуется перетяжка.
В итоге, получаются две дочерние клетки.
При реконструкции ядерной оболочки происходит сборка ядерных пор. Она начинается с образования ямки при слиянии внешней и внутренней ядерной мембраны, которая затем превращается в отверстие.
При сборке ядерной оболочки в телофазе белки ламины иммунохимически начинают выявляться в центромерных и теломерных участках хромосом, там же обнаруживаются первые признаки образования новой ядерной оболочки.
Судьба клеток
1. а) Часто образующиеся при делении дочерние клетки выходят из митотического цикла, т.е. далее не делятся. б) Таким образом они вступили в Go-период.
2. По этому признаку, т.е. по способности к делению, все клетки делятся на 3 типа. –
Первая категория
Постоянно делящиеся клетки.
Многие клетки организма, которые являются высокоспециализированными с течением времени изнашиваются и погибают.Они не способны к делению, однако они могут заменяться новыми. Эту функцию обеспечивают клетки того же типа, но которые не стали высокоспециализированными, и не утратили способность к делению. Эти клетки являются резервными.
-некоторые клетки базального слоя эпителия
-гемопоэтические клетки начальных стадий созревания (включая стволовые клетки).
Вторая категория
Неделящиеся клетки (в Go-периоде), сохранившие способность к делению при действии определённых стимулов.
В этом случае переход в Go-период обратим.
- клетки печени, а также
- стволовые клетки таких тканей как: (соединительные (фибробласты), костные (остеоциты), мышечные (ГМК) и т.п.), которые в обычных условиях не подвергаются постоянному клеточному обновлению.
Третья категория
Неделящиеся клетки (конечные), окончательно потерявшие способность делиться.
В данном случае переход из Go-периода необратим. Нет резерва для восстановления этих клеток
-нервные клетки,
-клетки сердечной мышцы.
Особенности митоза у растений и у животных
Растительная клетка Центриолей нет.
Звезды не образуются. Образуется клеточная пластинка. При цитокинезе не образуется борозды (перетяжки). Митозы происходят главным образом в меристемах.
Животная клетка Центриоли имеются
Звезды образуются. Клеточная пластинка не образуется При цитокинезе образуется борозда Митозы происходят в различных тканях и участках организма
АМИТОЗ
Это прямое деление интерфазного ядра путем перетяжки без образования хромосом вне митотического цикла. Амитоз может сопровождаться и делением клетки, но может делиться только ядро, что приводит к образованию многоядерных клеток. Может происходить даже сегментация ядра на несколько ядер. После амитоза клетки не в состоянии вступить в митотический цикл. Амитоз происходит только в узкоспециализированных клетках, которые больше не дадут начала новым типам клеток (например, клетки хрусталика глаза, клетки мочевого пузыря).
Для клеток это энергетически выгодно, так как хромосомы не образуются, не происходит их распределения, не формируется ахроматиновое веретено.
ЭНДОМИТОЗ
Удвоение числа хромосом в ядрах клеток многих растительных и некоторых животных организмов.
При эндомитозе не разрушаются ядерная оболочка и ядрышко, не образуется веретено деления клетки и не происходит реорганизация цитоплазмы, однако хромосомы проходят цикл спирализации и деспирализации.
Повторные эндомитозы приводят к возникновению гигантских полиплоидных ядер, отчего в клетке увеличивается содержание ДНК.
Эндомитоз это также процесс многократного удвоения нуклеопротеидных нитей - хромонем, составляющих основу хромосом, без увеличения числа последних; в результате образуются гигантские (политенные) хромосомы, что также связано со значительным увеличением в ядрах количества ДНК.
МЕЙОЗ
Биологическая сущность
Мейоз – это редукционное деление (деление созревания), потому что в результате мейоза у животных и некоторых растений образуются гаметы.
Мейоз – это особый способ деления, в ходе которого происходит уменьшение количества хромосом в дочерних клетках вдвое по сравнению с материнскими, т. е. происходит формирование гаплоидных клеток из диплоидных. Следовательно, мейоз обеспечивает чередование ядерных циклов (диплоидного – в организмах и гаплоидного – в гаметах), он гарантирует равновероятность и непрерывность передачи полного набора генетической информации в ряду поколений организмов в пределах биологического вида.
Основные события мейоза
Премейотическая интерфаза
Профаза I
Лептотена
Зиготена
Пахитена
Диплотена
Диакинез
Метафаза I
Анафаза
Телофаза
Интеркинез
Второе деление мейоза
Гаметогенез
Первое мейотическое деление
ПРОФАЗА I
лептотена (стадия тонких нитей) начинается спирализация хромосом.
зиготена (стадия сливающихся нитей),сближение и начало конъюгации гомологичных хромосом, которые объединяются в бивалент. Конъюгация сопровождается образованием лентоподобных структур между спаренными хромосомами - синаптонемальных комплексов (СК). В результате конъюгации образуются 23 тетрады (структуры из 4-х хроматид).
пахитена (стадия толстых нитей) между гомологичными хромосомами осуществляется кроссинговер
(Кроссинговер)
диплотена (стадия двойных нитей) отталкивание гомологичных хромосом, которые отделяются друг от друга в области центромер, но остаются связанными в областях прошедшего кроссинговера – хиазмах
диакинез (стадия обособления двойных нитей) гомологичные хромосомы удерживаются вместе лишь в отдельных точках хиазм
Метафаза I - завершается формирование веретена деления, его нити прикрепляются к центромерам хромосом, в результате чего биваленты устанавливаются в плоскости экватора веретена деления, образуя экваториальную пластинку.
Анафаза I - связи в бивалентах ослабляются и гомологичные хромосомы отходят друг от друга, направляясь к противоположным полюсам веретена деления. К каждому полюсу подходит гаплоидный набор хромосом, состоящий из двух хроматид.
Телофаза I - у полюсов веретена деления собирается одинарный гаплоидный набор хромосом, каждая из них содержит удвоенное количество ДНК (n2c).
Интеркинез - временной промежуток между первым и вторым делениями мейоза. Не всегда обязателен.
Мейоз 1
Интерфаза Практически отсутствует
Второе мейотическое отделение
(эквационное) протекает как митоз, только клетки, вступающие в него, несут гаплоидный набор хромосом.
В ходе второго деления хромосомы расходятся на хроматиды: образуются клетки - с гаплоидным содержанием и хромосом, и ДНК. При этом по виду полученной половой хромосомы мужские половые клетки разделяются на две группы - клетки с Х- и клетки с Y-хромосомой.
М ейоз
2
ейоз
2
В результате мейоза образуется
4 мужских клетки с гаплоидным набором хромосом и
1 женская клетка с гаплоидным набором хромосом
Ключевые особенности мейоза:
I. Конъюгация гомологичных хромосом и кроссинговер
На стадии профазы мейоза происходит конъюгация (попарное прилегание) гомологичных хромосом.
Кроссинговер - обмен хромосом строго гомологичными участками.
В результате, происходит рекомбинация (перераспределение) участков между гомологичными хромосомами.
II. Образование клеток с гаплоидным набором хромосом
Гаплоидные клетки образуются вследствие того, что в мейозе объединяются, по существу, два деления: первое - с предшествующим удвоением ДНК и второе - без удвоения ДНК.
В ходе первого деления тетрады расходятся на хромосомы, так что клетки получают по 23 хромосомы, состоящие из 2-х хроматид.
В ходе второго – клетки получают 23 хромосомы, но состоящие из 1-ой хроматиды
Старение клеток
Известный американский ученый Л.Хейфлик доказал, что естественная продолжительность жизни человека обусловлена числом митозов, которое могут совершить клетки данного организма. Оказалось, что клетки эмбриона могут совершить около 50 делений, а затем в них наблюдаются все признаки апоптотической смерти . У взрослого человека клетки могли совершить уже не 50 а гораздо меньше делений, в зависимости от возраста обследуемого пациента. Впоследствии было показано,что механизм старческого апоптоза запускается и находится в ядре.
Гибель клеток
Помимо процессов создания необходимого числа клеток в организме с помощью митоза существует и обратный процесс, который может приводить к уменьшению количества или к удалению старых, изношенных, поврежденных или просто лишних клеток
Апоптоз
генетически контролируемая гибель клеток. Будучи универсальным процессом элиминации клеток, апоптоз совместно с митозом обеспечивает регуляцию численности клеточных популяций. Апоптоз обеспечивает удаление клеток из нормально развивающихся и функционирующих тканей, не вызывая при этом повреждения соседних клеток и не запуская воспалительный процесс.
В начале процесса клетка утрачивает микроворсинки и контакты с соседними клетками, округляется и отделяется от клеточного пласта. Одновременно в ядре наблюдается перераспределение хроматина: он смещается к периферии, тогда как центральные области ядра становятся однородными. В результате гетерохроматин формирует по периметру ядра скопления с четко очерченными границами. Эухроматиновые области ядра при этом просветляются
Ядра клеток при апоптозе сжимаются, что обозначается термином “пикноз”.
Параллельно наблюдается конденсация цитоплазмы. При этом длительное время сохраняется целостность большинства цитоплазматических органелл, в том числе лизосом и митохондрий.
На более поздних этапах плазмолемма начинает формировать глубокие инвагинации, которые приводят к распаду клетки на гроздь апоптозных телец. В некоторых из них могут содержаться остатки клеточного ядра, состоящие из фрагментов нуклеолеммы и скоплений хроматина. В дальнейшем апоптозные тельца фагоцитируются макрофагами и другими клетками. Иногда апоптозные тельца не фагоцитируются, а слущиваются в полости, кровеносное русло или почечные канальцы. Длительность апоптоза обычно варьирует в пределах от 1 до 12 часов.
Некроз
Некроз - это патологический процесс, выражающийся в местной гибели ткани в живом организме в результате какого-либо экзо- или эндогенного ее повреждения. Некроз проявляется в набухании, денатурации и коагуляции цитоплазматических белков, разрушении клеточных органелл и, наконец, всей клетки. Некроз - это патологический процесс, запускающийся в ответ на какое-либо повреждающее воздействие (инфекция, химическое воздействие, облучение, недостаточное кровоснабжение и т.д.).
При некрозе происходит выход лизосомальных ферментов из лизосом, которые и переваривают содержимое клетки, клетка набухает и лопается. Содержимое клетки выбрасывается во внеклеточную среду, где поглощается фагоцитами, развивается воспаление. Существуют и другие формы программируемой гибели - аутофагия.
Процесс заключается в том, что органеллы соединяются с лизосомами, где перевариваются лизосомальными ферментами. Затем остатки клетки поглощают макрофаги
некроз |
апоптоз |
Морфологические свойства |
|
Имеется внешнее повреждающее клеточную мембрану воздействие |
Нет внешнего повреждающего воздействия |
Ядро сморщивается, хроматин конденсируется |
Ядро сморщивается,а хроматин конденсируется на конечных стадиях процесса |
Повреждаются внутриклеточные мембранные структуры |
Не повреждаются внутриклеточные мембранные структуры |
набухание и дезинтеграция органелл |
Органеллы остаются морфологически интактными |
Во внеклеточную среду выходят вещества обусловливающие воспалительную реакцию |
Не развивается воспалительная реакция |
Повреждаются митохондрии и нарушается энергообразование |
Нарушается энергообразование в митохондриях |
Биохимические свойства |
|
потеря регуляции ионного гомеостаза |
Регулируемый процесс, включающий активацию и энзиматические реакции |
Не требует энергии – пассивный процесс, может происходить при температуре +4С |
Энергозависимый процесс – активный, требует физиологической температуры |
Расщепление ДНК в случайных точках (большое размытое пятно ДНК при электрофорезе |
Специфическое расщепление ДНК до моно- и олигонуклеотидов (лестница ДНК при электрофорезе в агарозном геле) |
Физиологическое значение |
|
Отмирание одиночной клетки |
Отмирание группы клеток |
Клетка разрушается и ее части поглощаются иммунокомпетентными клетками |
Элементы цитоплазмы отщепляются в мембранных пузырьках и поглощаются соседними клетками или тканевыми макрофагами |
На месте погибшей клетки происходит соединительнотканое замещение |
На месте погибшей клетки не происходит соединительнотканое замещение |
Регенерация тканей
Регенерацией называется образование новой ткани на месте погибшей, отмершей. В здоровом, нормальном организме все время происходит физиологическая регенерация клеток; постоянно слущивается отмерший роговой слой эпидермиса, и взамен него во внутреннем слое кожи размножаются новые клетки. Такое же слущивание покровного эпителия происходит и на слизистых оболочках. В кровеносных сосудах эритроциты обычно живут 60—120 дней. Следовательно, приблизительно в течение 2 месяцев происходит полное их обновление. Так же систематически восполняются по мере их гибели или отмирания и лейкоциты, и другие форменные элементы крови. При различных патологических процессах клетки и ткани разрушаются в большем количестве, чем в норме. Регенерации тканей принадлежит огромное значение в процессе восстановления поврежденных тканей и органов. В регенерации различают такие понятия, как форма регенерации, уровень регенерации, способ регенерации.
Формы регенерации:
1. Физиологическая регенерация - восстановление клеток ткани после их естественной гибели (например, кроветворение);
2. Репаративная регенерация - восстановление тканей и органов после их повреждения (травмы, воспаления, хирургического воздействия и так далее).
Регуляция регенерации тканей
Регенерация тканей происходит вследствие пролиферации недифференцированных клеток, обладающих способностью не только делится под действием соответствующих стимулов, но также и дифференцироваться в клетки той ткани, регенерация которой происходит. Эти клетки носят название взрослых стволовых клеток. Многие ткани взрослого организма, такие как ткани гемопоэтической системы, пищеварительный эпителий, мозг, эпидермис, легкие содержат пул таких клеток. Стволовые клетки тканей взрослого индивидуума снабжают организм зрелыми дифференцированными клетками в течение нормального гомеостаза, а также во время регенерации и восстановления тканей и органов.
Стволовая клетка
По широте дифференцировочного потенциала СК делят на:
- тотипотентные - способны полностью воссоздавать ткани и органы всего организма; яйцеклетка (зигота), а также бластомеры;
тотипотентность зиготы обеспечивает развитие как самого эмбриона, так и внезародышевых органов и тканей - плаценты, амниотического мешка, пуповины),
плюрипотентные - формируют ткани всех трех зародышевых листков – эктодермы, эндодермы и мезодермы; (ЭСК из внутренней клеточной массы бластоцисты),
-мультипотентные-способны дифференциро-ваться в специализированные клетки нескольких типов в пределах одного зародышевого листка.
олигопотентные и бипотентные СК, направления дифференцировки которых достаточно ограничены.
- унипотентные - дифференцируются только в специализированные клетки одного определенного типа, (сперматогониальные СК).