Волосы

Волосы характерны лишь для представителей класса млекопитающих, которых в связи с этим называют еще как Trichozoa (волосатые животные), или Pelifera (несущие волосы). Наиболее древние находки подобных волосам структур имеют возраст 200–210 млн лет. Уже у вероятных предков млекопитающих – териодонтов (Theriodontia) на конце морды и по всей верхней губе имелось сенсорное поле и, возможно, присутствовали «протовибриссы».
С уществуют
гипотезы, согласно которым волосы
произошли от роговых осязательных
волосков чешуйчатых пресмыкающихся,
или прототрихий («предшественников
волос») – бесхвостых земноводных
(которые действительно очень напоминают
эмбриональные волосы), или органов
боковой линии личинок водных хвостатых
земноводных, которые могли соответствующим
образом видоизмениться в связи с выходом
на сушу.
уществуют
гипотезы, согласно которым волосы
произошли от роговых осязательных
волосков чешуйчатых пресмыкающихся,
или прототрихий («предшественников
волос») – бесхвостых земноводных
(которые действительно очень напоминают
эмбриональные волосы), или органов
боковой линии личинок водных хвостатых
земноводных, которые могли соответствующим
образом видоизмениться в связи с выходом
на сушу.
Действительно, у зародышей млекопитающих (и человека в том числе) развитию чешуй и волос предшествует увеличение числа чешуевидных клеток в покровах, что иногда считают кратковременным эмбриональным повторением стадий развития органа предков. Существует заболевание «черный волосатый язык», при котором у человека нитевидные сосочки языка гипертрофируются и приобретают вид настоящих волос – т.е. в принципе такое видоизменение роговых структур возможно.
О днако
в настоящее время превалирует мнение,
что волосы – новоприобретение
млекопитающих и эволюционно не связаны
с какими-либо производными наружных
покровов их предков. Становление волос,
возможно, связано с развитием кожных
желез. Ороговение эпидермиса – секреторный
процесс; чешуи, перья и волосы являются
железистыми структурами, секрет которых
– кератин – выполняет защитную функцию.
днако
в настоящее время превалирует мнение,
что волосы – новоприобретение
млекопитающих и эволюционно не связаны
с какими-либо производными наружных
покровов их предков. Становление волос,
возможно, связано с развитием кожных
желез. Ороговение эпидермиса – секреторный
процесс; чешуи, перья и волосы являются
железистыми структурами, секрет которых
– кератин – выполняет защитную функцию.
Терапсида (Therapsida – зверообразные пресмыкающиеся) были водными и полуводными формами с мягкой эластичной и, видимо, железистой кожей, которая в процессе их эволюционного выхода на сушу ороговевала и становилась влагонепроницаемой. Железы опускались глубоко в кожу, изолируясь от кератиновых структур, а последние превратились в волосы и чешуи.
Непосредственными предками млекопитающих были, вероятно, мелкие хищные рептилии цинодонты (Cynodontia). В их различных семействах и родах тем или иным образом сочетались признаки как пресмыкающихся, так и млекопитающих. Предполагают, что по крайней мере наиболее эволюционно продвинутые представители цинодонтов обладали такими особенностями зверей, как теплокровность и выработка молока для питания детенышей. Развитие шерстного покрова при этом имело теплоизолирующее значение и было важным для становления гомойотермии. Впрочем имеется мнение, что первоначальной функцией волосяного покрова у древних млекопитающих была, напротив, защита от перегрева, так как неспособность интенсивно отдавать тепло тормозила эволюцию класса в условиях жаркого и влажного климата мезозоя.
Волосы, вероятно, имелись уже у потомков цинодонтов (230 млн лет назад), причем у ветви, давшей яйцекладущих млекопитающих, это были только волосы как таковые, а у ветви, приведшей к возникновению сумчатых и плацентарных, – и волосы и вибриссы (чувствующие волосы). У современных однопроходных вибрисс нет (у них развиты кожные механо- и электрорецепторы), у сумчатых вибриссы присутствуют на морде и конечностях, а у плацентарных их распространение на теле может быть самым разнообразным.
Качественные и количественные различия между собственно волосами и вибриссами настолько велики, что ряд исследователей задаются вопросом: а волосы ли вибриссы? Движение вибрисс обеспечивается поперечнополосатой мускулатурой, волос – гладкой (единственное исключение или, возможно, одна из параллельных ветвей развития – волосы однопроходных, которые снабжены поперечнополосатой мускулатурой). Соединительнотканная капсула у вибриссной сумки развита значительно сильнее, чем у волосяной. Вибриссы не имеют потовых желез, а сопутствующие сальные железы развиты слабо. Различия между волосами и вибриссами касаются также характера их кровоснабжения и иннервации, строения (у вибрисс отсутствует сердцевинный слой), характера линьки (вибриссы сменяются по одной по мере стирания, независимо от общей линьки), принципа функционирования (вибрисса как подвижный рычаг) и самого их функционального значения (чувствительного у вибрисс и теплоизолирующего у волос).

Строение волоса (слева) и вибриссы (справа)
Косвенными доказательствами возможного разделения путей развития волос и вибрисс служат также опережающее развитие иннервации еще не сформированного фолликула вибриссы в онтогенезе, возможность обратного развития вибрисс в рецепторы (как это происходит у дельфинов), глубокое залегание фолликулов вибрисс в подкожной клетчатке и обособление их мышц от подкожной поперечнополосатой мускулатуры и, наконец, представительство каждой вибриссы в сенсорных зонах мозга.
Вибриссы могут либо двигаться волевым усилием («активные»), либо непроизвольно, либо вообще не двигаться («пассивные»). У человекообразных обезьян редкие (2–3 пучка) лицевые вибриссы не имеют собственных мышц и приводятся в движение мимической мускулатурой; у лошадиных, быков и представителей семейства свиных они неподвижны.
У грызунов вибриссы снабжены крупными мышцами и активно двигаются.
|
|


При общей редукции волосяного покрова вибриссы сохраняются и даже могут равномерно распределяться по телу – как у сиреновых.
На голове у китов полосатиков (Balaenoptera) присутствуют многочисленные вибриссы – не рудиментарные органы некогда хорошо развитого волосяного покрова, а вполне специализированные органы чувств. У зубатых китов вибриссы имеются только у эмбрионов. У взрослых особей большинства представителей семейства дельфиновых (Delphinidae) в коже морды остаются «ямки» после дегенерации ювенильных вибрисс, и, как уже было сказано, возможно, происходит инволюция их фолликулов в специализированные тактильные органы, реагирующие на изменение давления воды и низкочастотные колебания.
Можно предположить, что у предковых форм млекопитающих вибриссы возникли из эмбриональных закладок примитивных механорецепторов, в то время как волосы – позднее – от чешуевидных эпидермальных закладок.
Жесткий прямой стержень вибрисс и отсутствие у них характерного для волос ворса свидетельствуют, что они появились первыми – у таких предковых форм, у которых толстая шкура не была прочно прикреплена к туловищу. Волосы же возникли у более поздних форм, у которых, по мере оформления подкожных структур, кожа стала тоньше, но прочнее соединялась с туловищем, и растущие волосы приобрели наклон вследствие натягивания, растягивания и роста такой закрепленной кожи. Этот процесс прослеживается и в индивидуальном развитии: первыми, на этапе еще окончательно не оформленных и не прикрепленных к подкожной мускулатуре слоев кожи, закладываются вибриссы, а волосы – позднее, в полностью сформированной, более тонкой и прикрепленной коже. У зрелорождающихся форм вибриссы в процессе индивидуального развития закладываются раньше, но развиваются медленнее и оформляются в более поздний эмбриональный период, чем у незрелорождающихся.
|
Американский дикобраз (слева): 1 – вершина иглы-гарпуна; 2 – поперечный срез через сердцевину иглы Пятнистый олень (вверху): 1 – кутикула основания остевого волоса; 2 – кутикула гранны; 3 – пуховой волос; 4 – поперечный срез через сердцевину остевого волоса |


Несмотря на различия как вибриссы, так и все другие типы волос и их производных (иглы, полуиглы, колючки, полуколючки, щетины, кроющие, направляющие, остевые разных категорий, пуховые, тилотрихии, монотрихии, осметрихии и др.) имеют общий тип микроструктуры. Стержень волоса – ороговевший цилиндр различной конфигурации, покрытый одно- или многослойной кутикулой, заполненный плотными веретеновидными корковыми клетками и не всегда присутствующим центральным нежным сердцевинным слоем – ороговевшими остатками стенок сердцевинных клеток метаболически активного фолликула.
С троение
волосяной луковицы:
1 – наружное
корневое влагалище;
2 – слой Генли;
3
– слой Гексли;
4 – кутикула внутреннего
корневого влагалища;
5 – сердцевина;
6
– кора;
7 – кутикула коры
троение
волосяной луковицы:
1 – наружное
корневое влагалище;
2 – слой Генли;
3
– слой Гексли;
4 – кутикула внутреннего
корневого влагалища;
5 – сердцевина;
6
– кора;
7 – кутикула коры
Волосяной фолликул – динамичная комплексная система, подверженная возрастным и сезонным изменениям под гормональным контролем, с достаточно большой функциональной пластичностью, позволяющей изменять параметры волоса. Считается, что фолликулы возникают лишь единожды в онтогенезе, однако экспериментально доказано новообразование волос при регенерации кожи.
При индивидуальном развитии волосяного покрова имеет место гетерохрония – разница во времени закладки и развития различных типов волос. Сначала, как уже сказано, формируются самые сложные структуры – вибриссы, затем чувствующие волосы – снабженные капиллярами и нервами тилотрихии, а еще позже – волосы разных типов – от сложных остевых (из первичных фолликулов, снабжены мышцами и железами) полуигл, игл, щетин до просто устроенных пуховых (из самостоятельных закладок или из вторичных фолликулов). Первичные фолликулы происходят из эпидермиса, а вторичные – продукт почкования первичных.
Весьма интересен процесс эволюционного становления игл у млекопитающих. Образование игл происходило в разных группах параллельно и в связи с конкретными приспособлениями к образу жизни. О том, что иглы – модификация волос, свидетельствуют многочисленные переходные формы (щетины, полуиглы). У эмбрионов ежа иглы закладываются позднее волос и образуются путем слияния нескольких фолликулов, что довольно обычно у видов, у которых имеются пучки волос, растущие из общего фолликула. Однако образование иглы у ежа – не механическое объединение фолликулов, а более сложный процесс. Каждая игла формируется в результате разной скорости развития группы фолликулов, в которой центральный, самый крупный, фолликул развивается быстрее других и, разрастаясь, «захватывает» более мелкие и медленнее растущие. В итоге игла оказывается состоящей из разделенных толстыми продольными перегородками тяжей сердцевинных клеток, среди которых центральный тяж наиболее крупный. У представителей совсем другой группы – дикобразов – иглы устроены проще: перегородки гораздо тоньше, а сердцевинные клетки либо одинаковы и по форме и по размерам, либо немного крупнее в центральной части стержня. В бороздчатых иглах и полуиглах некоторых хомяковых и мышиных (разумеется, у тех представителей этих семейств, у которых такие образования имеются) сердцевина сохраняется лишь в боковых частях иглы и разделена утолщенным корковым слоем, что предполагает формирование этих иголок из одной крупной закладки волоса без сердцевины и двух более мелких – с сердцевиной. Щетины свиных и пекариевых (Tayassuidae) очень похожи на «комбинированные» иглы и также состоят из разделенных толстыми перегородками тяжей сердцевинных клеток. В иглах (или полуиглах) тенреков и ехидн продольные перегородки отсутствуют, сердцевинные клетки имеют сходные размеры и отличаются утолщенными стенками и слабым развитием полостей – т.е. эти структуры формируются из единой закладки.
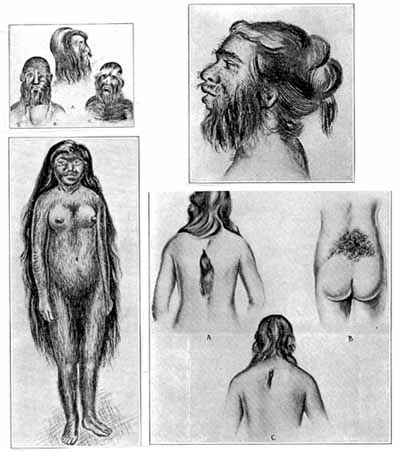
Редукцию волос у человека связывают со становлением бипедии – двуногости, фетализацией – замедлением темпов развития в онтогенезе отдельных органов и их частей и половым отбором – предпочтением безволосых половых партнеров и своеобразной сексуальной революции – получения наслаждения при любовных играх.
Возможно, роль важного фактора сыграл тепловой стресс – при формировании адаптивного типа конституции у ранних тропических гоминид основным механизмом терморегуляции стало интенсивное потоотделение. Волосяной покров затруднял отдачу тепла и был утерян.
Замедленное развитие под гормональным контролем приводит к подавлению ряда признаков, в частности волосяного покрова, т.е. с определенной точки зрения человека можно рассматривать как зародыша примата, достигшего половой зрелости (своеобразная неотения).
Как и у человека, выношенные плоды гориллы и шимпанзе имеют длинные волосы на голове, но слабо оволосненные морду и туловище, а облысение у них проходит сходно с таковым у человека.
Эмбриональные волосы (vellus, лануго) человека развиваются раньше более грубых и крупных «терминальных» волос. Закладки волос появляются в разное время на лице и голове, затем по всему телу сверху вниз. Считают, что волосы плода человека – рекапитуляция (повторение у эмбрионов признаков взрослых предков) волос человекообразных обезьян, однако показана и их функциональная значимость: реакция этих волос в ответ на движение околоплодных вод (ворс направлен против него, и волосы действуют как рычаги с длинным плечом) усиливает деятельность кожных рецепторов и поддерживает мышечный тонус, обеспечивающий определенную позу плода.
При гормональной недостаточности или вследствие генных мутаций у человека могут развиваться патологические состояния, например, чрезмерная волосатость – гипертрихоз. В этом случае зародыш покрыт длинными, тонкими, шелковистыми и волнистыми, как у болонки, волосами («люди-собаки»), которые растут и остаются на всю жизнь (волосатый человек Андриан Евстихиев).