

18. Механизмы переноса электрона и миграции энергии в биоструктурах.
Молекулярные механизмы биологических процессов во многом обусловлены электронными свойствами молекул. Изменение электронного состояния одного участка (активный центр) дает начало последовательным конформационным превращениям, захватывающим большие области в макромолекуле. Рассмотрим физические механизмы процессов трансформации электронной энергии в биоструктурах. Особое значение имеют миграция энергии электронного возмущения и транспорт электронов.
Транспорт электрона может происходить на относительно большие расстояния и независимо от поступательных движений донора и акцептора электрона.
Рассмотрим концепцию туннельного транспорта электрона между отдельными белковыми молекулами – переносчиками, отделенными друг от друга энергетическими барьерами.
Туннельный транспорт происходит в условиях, когда значение энергии электрона меньше высоты энергетического барьера между молекулами.
Осуществляется на расстояниях порядка 0,5 нм.
Протекает при низких температурах – 77 К и ниже.
Перенос электрона происходит в фотосинтетических реакционных центрах, зависит от конформационного состояния белковых компонентов (рис. 3.22).
Туннельный перенос электрона имеет место и в митохондриях, также сопряжен с конформационными изменениями белков.
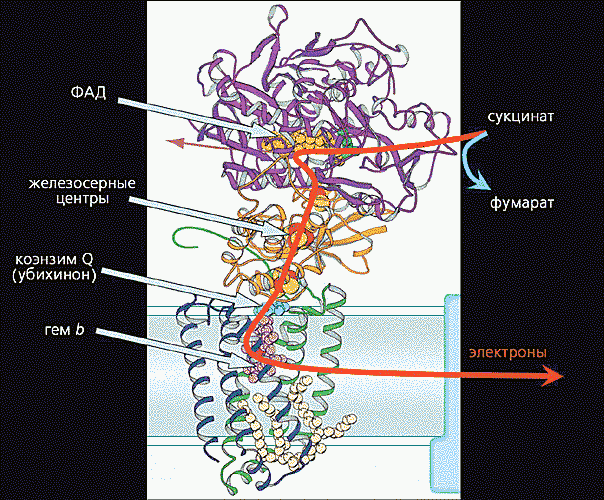
Рис. 3.22. Туннельный перенос электрона в митохондрии
В результате возмущения молекулы донора или присоединения к ней электрона от внешнего источника система оказывается в возмущенном состоянии.
Необратимый перенос энергии или электрона от донора к акцептору возможен лишь в случае рассеивания части энергии возбуждения (электрона) за время пребывания в акцепторе.
На фоне квантово-механических колебаний между двумя электронными состояниями (i – f) вероятность перехода между состояниями ψi и ψf в единицу времени Wif, определяется формулой («золотое» правило Ферми):
Wif = 2π |Vif|2 ρf (Ei) / ħ,
ρf – плотность уровней в конечном состоянии при энергии, равной Ei ,
Vif – электронный матричный элемент, определяется энергией кулоновского взаимодействия между донором и акцептором и перекрыванием электронных волновых функций донора и акцептора;
ħ = h/2π (h – постоянная Планка).
Перенос электрона между белковыми макромолекулами связан с конкретными путями движения электрона внутри отдельных молекул, а также с образованием контактных комплексов на их поверхностях. В белке туннелирование электрона между донором и акцептором осуществляется с участием виртуальных уровней промежуточных звеньев аминокислотной последовательности, которые играют роль электронных «мостиков». Электронные «мостики» зависят от конформационного состояния глобулы, в переносе участвуют различные группы и связи между ними.
Электронно-конформационные взаимодействия включают собственно перенос электрона и внутримолекулярное движение в белке по конформационным подсостояниям. Скорость переноса электрона зависит от конформации определенных фрагментов цепи.
При поглощении кванта света меняется баланс сил, определяющий равновесную конформацию донорно-акцепторной пары в темноте. Осуществляется фотоконформационный переход. Изменяются параметры, определяющие вероятность туннелирования электрона. В фотосинтетической системе переноса электрона они включают образование таких контактных состояний между переносчиками, которые обеспечивают направленное туннелирование между ними в транспортной цепи.
Индуктивно-резонансный механизм миграции энергии.
Перенос энергии происходит за счет кулоновского взаимодействия между электронами в молекулах Д и А. Во время существования возбужденного состояния молекулы Д* генерируется переменное электромагнитное поле за счет осцилляции заряда электрона.
Это поле взаимодействует с электроном в невозбужденной молекуле акцептора А. Если частота переменного поля Д* совпадает с частотой перехода электрона в А на возбужденный уровень, то происходит перенос энергии.
При этом молекула Д* возвращается в основное, а молекула А переходит в возбужденное состояние. Никакого высвечивания кванта света молекулой Д* и перепоглощения его молекулой А не происходит, а перенос носит безызлучательный характер.
Индуктивно-резонансный перенос возможен не только между синглетными уровнями донора и акцептора, но и по триплет-синглетному и синглет-триплетному типу.
Обменно-резонансный механизм миграции энергии возможен на более коротких расстояниях, когда электронные орбитали донора и акцептора могут перекрываться. Тогда происходит "обмен" электронами и электронными состояниями.
Экситонный механизм миграции энергии осуществляется при больших энергиях взаимодействия.
Возбуждение, попавшее в молекулу донора, может перейти в соседнюю молекулу акцептора раньше, чем успеет произойти релаксация на нижние колебательные уровни состояния молекулы донора.
Возбуждение как бы бежит по верхним колебательным уровням взаимодействующих молекул, не успевая локализоваться на каждой из них в отдельности.
Возбуждением одновременно охвачено несколько сот молекул, и оно носит коллективный характер. Область возбуждения, включающая большое число молекул, называется экситоном.
В фотосинтетических мембранах экситонный механизм имеет место при миграции энергии в пределах группы однородных молекул пигмента.