

58 Рецепторы. Рецепторный и генераторный потенциал.
Р -ы
представл. собой специализир. образования,
воспринимающие определенные виды
раздражений. Р.обладают наибольшей
чувствительностью к адекватным для них
раздражениям. Р. делят на 4 группы:
механо-,термо-, хемо- и фоторец-ры. Каждую
группу подразделяют на более узкие
диапазоны рецепции. Например, зрительные
рец-ры деляться на воспринимающие
освещенность, цвет, слуховые- определ.
тон, вкусовые- опреднл. вкусов. раздражения.
Рец-ый потен-л, возникает при раздражении
рец-ра как результат деполяризации и
повышения проводимости участка его
мембраны, который называется реце-ым.
Рецептивный участок имеет спицифические
свойства, в том числе и биохимические,
отлич. его от мембраны тела и аксона.
Возникший в рецептивных участках
мембраны рец. потенциал электротонически
распространяется на аксонный холмик
рец. нейрона, где возникает генераторный
потенциал. Его возникновение в области
аксонного холмика объясняется тем, что
этот участок нейрона имеет более низкие
пороги возбуждения и ПД в нем развивается
раньше, чем в других частях мембраны.
Чем выше ген. пот-л, тем интенсивнее
частота разрядов распространяющегося
ПД от аксона к другим отделам нервной
системы. Значит, частота разрядов
рецепторного нейрона зависит от амплитуды
ген-го пот-ла. Рец-ые нейроны различ-ся
по скорости уменьшения их реакции на
длящуюся стимуляцию. Рец-ые ней-ны
медленно адаптир-ся к раздр-ию, т.е.
длительное время генерирующие ПД наз
тоническими. Рец-ры, быстро и скоро
реагирующие на стимуляцию группой
импульсов, наз фазическими. Таким
образом, реакция рецепторного нейрона
предназначенного для передачи инф-ии
из области восприятия имеет 5 стадий 1.
преобразование сигнала внешнего
раздражения 2. генерация рец-го пот-ла
3. распрастранение рец-го пот-ла по
нейрону 4. возникновение генер-го пот-ла
5. генерация нер-го импульса.
-ы
представл. собой специализир. образования,
воспринимающие определенные виды
раздражений. Р.обладают наибольшей
чувствительностью к адекватным для них
раздражениям. Р. делят на 4 группы:
механо-,термо-, хемо- и фоторец-ры. Каждую
группу подразделяют на более узкие
диапазоны рецепции. Например, зрительные
рец-ры деляться на воспринимающие
освещенность, цвет, слуховые- определ.
тон, вкусовые- опреднл. вкусов. раздражения.
Рец-ый потен-л, возникает при раздражении
рец-ра как результат деполяризации и
повышения проводимости участка его
мембраны, который называется реце-ым.
Рецептивный участок имеет спицифические
свойства, в том числе и биохимические,
отлич. его от мембраны тела и аксона.
Возникший в рецептивных участках
мембраны рец. потенциал электротонически
распространяется на аксонный холмик
рец. нейрона, где возникает генераторный
потенциал. Его возникновение в области
аксонного холмика объясняется тем, что
этот участок нейрона имеет более низкие
пороги возбуждения и ПД в нем развивается
раньше, чем в других частях мембраны.
Чем выше ген. пот-л, тем интенсивнее
частота разрядов распространяющегося
ПД от аксона к другим отделам нервной
системы. Значит, частота разрядов
рецепторного нейрона зависит от амплитуды
ген-го пот-ла. Рец-ые нейроны различ-ся
по скорости уменьшения их реакции на
длящуюся стимуляцию. Рец-ые ней-ны
медленно адаптир-ся к раздр-ию, т.е.
длительное время генерирующие ПД наз
тоническими. Рец-ры, быстро и скоро
реагирующие на стимуляцию группой
импульсов, наз фазическими. Таким
образом, реакция рецепторного нейрона
предназначенного для передачи инф-ии
из области восприятия имеет 5 стадий 1.
преобразование сигнала внешнего
раздражения 2. генерация рец-го пот-ла
3. распрастранение рец-го пот-ла по
нейрону 4. возникновение генер-го пот-ла
5. генерация нер-го импульса.
Общий биофизический механизм восприятия рецепторами и клетками раздражений.
Обработка сенсорной информации начинается в специальных мембранах рецепторных клеток. Рецепторные клетки обладают двумя важными свойствами:
1.Высокая избирательность по отношению к стимулам определенной модальности (т.е. обладает дифференц. чувствительностью и преобразуют в биол. сигналы лишь определенные формы энергии, что связано с конформационным изменением рецепторной молекулы). Активация рецепт. мол-лы вызывает в рецепторах разных типов разную последовательность молекулярных событий. В некоторых рецепторах активация рецепт. мол-лы инициирует каскад ферментативных реакций, служащий для усиления сигнала. При этом каждая последующая реакция сопровождается многократным (10-1000 раз) увеличением числа активных молекул, за счет чего достигается усиление сигнала. Последняя реакция каскада приводит к открытию или закрытию ионных каналов, которые проводят рецепторный ток, модулирующий нервный сигнал. Т.о., рецепторная клетка преобразует внешний стимул в трансмембранный ток, вызывающий изменение мембранного потенциала. Преобразование стимула и избирательность рецепторных клеток в основном определяется чувствительными молекулами, находящимися в клеточной мембране рецептора.
2.Способность генерировать электрический сигнал, энергия которого намного больше энергии внешнего стимула.

Рецепторный белок либо сам может быть частью ионного канала, либо может модулировать активность мембранных каналов косвенным образом через каскад ферментативных реакций, усиливающий клеточный сигнал.
61 Локализация гена.
Локализовать – значит установить место гена в геноме.
Этот процесс связан с построением генетической карты. Методов локализации оч. много:
1.Сцепление с X или Y – хромосомами устанавливают по картине наследования.
2.Метод генетических маркеров (тестеров) – локализация в аутосомах. Используются маркеры из установленных групп сцепления.
Дробный метод рецессивно – генетических маркеров.
Предлагается 2 этапа (если не сцеплено с полом):
установление группы сцепления (аутосомный). Необходимое условие – наличие линий тестеров в каждой группе сцепления. Необходимо иметь хотя бы по одному маркеру.
Условие – наличие не менее 2ух партнеров из той группы сцепления, в которую попал локализуемый ген.
3. Цитогенетические методы: - совместно с использованием генетических карт используются нуллисомики, моносомики, трисомики + маркер; - хромосомные перестройки (делеции, дупликации, инверсии и т.д.)
4. Методы клеточной биологии. При совместном культивировании кл. разных видов, происходит соматическая гибридизация, а далее утеря хромосом одного вида. Выбирают клоны, у которых наряду со своими хромосомами есть одна хромосома от другого вида. Метод сочетается с дифференциальным окрашиванием и биохимическими методами => виды должны отличатся по биохимическим маркерам. По анализу электрофоретической подвижности ферментов (имеется в норме у донора хромосомы) делают вывод. Если эти характеристики соотв-ют тем, что в норме есть у донора данной хромосомы, то делают вывод о том, ген этого фермента локализован в данной хромосоме, далее используют транслокации.
5. Молекулярно - генетические методы. Связаны с гибридизацией нуклеиновых кислот in situ.
6. генеалогический метод для генов человека
7. трансдукция и трансформация у бактерий
Теория гена. Эволюция представлений о гене, современные представления о строении гена.
В настоящее время ген определяют как структурную единицу генетической информации, далее неделимую в функциональном отношении. Ген представлен участком молекулы ДНК (реже РНК).
Проблема гена – центральная проблема генетики. Представления о гене всегда отражали в концентрированной форме уровень развития, достижения и нерешенные проблемы генетики. Понятие «ген» как дискретной единицы, выявляемой менделевским гибридологическим анализом, ввел В.Л. Иоганнсен в 1909 году. Он не связывал это понятие с какими-либо гипотезами о его сущности и материальной природе.
«Слово «ген» совершенно свободно от какой бы то ни было гипотезы; оно выражает лишь тот точно установленный факт, что многие признаки организма обуславливаются в гаметах особыми, отделимыми и потому самостоятельными «состояниями», «основами», «задатками», короче – тем, что мы будем называть «генами», - писал Иоганнсен. Такое представление о гене было характерно для периода классической, или формальной, генетики.
С тех пор произошла не только «материализация» гена, но сами гены - участки молекул ДНК – стали объектами и рабочими инструментами генной инженерии и биотехнологии.
Расшифрована первичная структура тысяч генов, выяснены основные черты и разнообразие их строение у разных объектов. Все эти сведения хранятся в компьютерных банках информации, используемых и пополняемых учеными всего мира.
Первая успешная попытка конкретизации представлений о гене принадлежит Т.Х. Моргану, который один из своих классических трудов назвал «Теория гена» (1926 год). Согласно его теории гены находятся в хромосомах и представляют собой далее не делимые единицы мутации, рекомбинации и функции. Ген – это:
Единица мутации, то есть ген изменяется как целое
Единица рекомбинации, то есть кроссинговер никогда не наблюдали в пределах гена
Единица функции, то есть все мутации одного гена нарушают одну и ту же генетическую функцию, что выражается в их некомплиментарности у особей F1 при попарном скрещивании мутантов
Современные представления:
Гены закреплены на внутренней оболочки клеточного ядра и находятся в ядерном матриксе, образуя петлеобразную структуру. Длина щели между ними от 20-200 пар нктд. В одной петле может располагаться несколько генов. Энхансеры регулируют последовательность включения этих генов в развитии организма. Положение гена в петле не случайно: чем ближе к энхансеру, тем эффективнее ген. Петли разграничены инсуляторами, взаимодействующими со специфическими белками. Функцию инсулятора выполняет нормальная последовательность ДНК, связанная со специфическими белками.
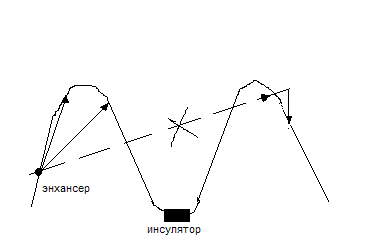
если убрать инсулятор, то энхансер будет контролировать и прерывистое направление гена. Инсуляторы обеспечивают независимость гена от других последовательностей в геноме. Раньше инсулятор называли НКЛ – последовательность, контролирующая локус. Эта последовательность содержит сильные энхансеры и сайты прикрепления к ядерному матриксу. Роль последовательностей между сайленсорами и энхансерами неизвестна. Структура этих участков вариабельна.
Эукариотический ген – единица транскрипции, обеспечивающая синтез одного полипептида или набора близкородственных полипептидных цепей.
Генетический локус – ген и цис-регуляторные элементы.
Не всегда ген кодирует полипептид, ген может отвечать только за синтез РНК, но определение гена, как единицы транскрипции, сохраняется.
Единица транскрипции может кодировать более одного белка.
Известно формирование нескольких генов на базе одной и той же полинуклеотидной цепи.
Кодирующие последовательности могут располагаться в разных цепях (в геномах вирусов).
В некоторых случаях регуляторные элементы (энхансер, сайленсор, промотор) располагаются внутри единицы транскрипции. В этом случае (тРНК, 5S рРНК и мяРНК) понятие гена и генетического локуса совпадают. У эукариот огромное многообразие путей эволюции структурной организации генов, что позволяет говорить о значительном усложнении регуляции генома в целом.