

2.2. Потік енергії через популяцію
Особина впродовж активного життя непомітно пов'язана з середовищем якраз потоком енергії і матерії, котрий надходить разом з кормом із середовища до особини, в якій ці два компоненти і матерія нагромаджуються у вигляді біомаси. Одночасно частина енергії та матерії після певних перетворень особиною надходить до середовища у вигляді продуктів обміну речовин і тепла. Цей процес має назву біологічної продуктивності організму. Той самий процес, що й у випадку однієї особини, вирішує долю життя, приросту біомаси, народжуваності і тривалості життя конкретної популяції рослин чи тварин. Потік енергії і матерії через популяцію називають біологічною продукцією популяції.
Для того, щоб створити нову матерію – продукцію, необхідний корм (С). Наступний процес засвоєння особиною частини матерії у вигляді корму становить брутто продукції, або ж асиміляції (А). Частина засвоєної продукції йде на побудову тіла особини, яке називають біомасою (продукція нетто Р). Наявна в кормі енергія використовується на життєві процеси особини (пересування, втеча, погоня, пережовування корму і т.п.), а також розсіюється у вигляді теплової, хімічної (наприклад, газовиділення), світлової, електричної енергії. Оскільки всі організми беруть енергію з окисленої органічної субстанції (вуглецю), її мірилом є інтенсивність дихання особин (респірація (R), від лат. "респіро" – дихання).
У тварин неперетравлений корм, а також фекальні рештки (кал, сеча або урина), які виділяються в середовище (FU), є не що інше, як різниця між спожитою їжею і асиміляцією (FU=(C-A)).
Подібно до тварин рослини виділяють корінням і надземними органами продукти асиміляції: вуглекислий газ, токсичні метаболіти, а також невикористані метаболізмом сполуки. Як ми вже знаємо, ці виділення можуть бути для інших організмів шкідливими (алелопатія), корисними (біостимуляція) або ж можуть не приносити ні шкоди, ні користі. Крім згаданих виділень, які утворюються внаслідок засвоєння корму, тварини і рослини виділяють ще різноманітні слизи. Всі вони разом (фекалії, сеча, слиз і т.п.) і будуть становити непродуктивні відходи життєдіяльності особин (FU).
Розглянемо загальні рівняння енергетичного балансу для особин і популяції, сформульовані польським біологом К.Петрусевичем (1978):
С = Р + R + FU;
А = Р + R = С - FU;
P = A - R = C - FU - R.
Однак слід зауважити, що ці рівняння енергетичного балансу певною мірою спрощені і мають теоретичний характер. Якщо б навіть вдалося в лабораторних умовах контролювати всі названі параметри, навряд чи можна було б домогтися такого ідеального балансу [1].
2.3. Продуктивність популяції
Розглядаючи ідею продуктивності в екології, К. Петрусевич наводить такий приклад. Якщо в певний момент Т1, маємо, наприклад, 10 особин (N1), а в момент Т2 їх виявилося N2=20 особин, то це означає, що різниця N2 - N1 = ΔN = 10. Це може бути у випадку народження 10 особин і відсутності відпаду або у випадку народження 25 особин і загибелі 15, а також при народженні 100 особин і загибелі 90.
Таким чином, різниця стану, підкреслює К. Петрусевич, говорить нам дуже мало або й зовсім нічого про те, який був доплив особин до популяції. Отже, збільшення особин у популяції і є її продукцією, вираженою в особинах. Однак кожна особина має свою масу, а отже, сумарна маса особин популяції являє собою масу популяції, яка може виражатися в одиницях маси – тоннах, центнерах, кілограмах, грамах чи міліграмах.
У першому і другому випадках у енергетичному балансі особин продукція складається з двох елементів: приросту чи зменшення біомаси і одночасно продукції потомства. Причини спаду динаміки є різні: натуральна смертність, втрати, які несе популяція внаслідок її експлуатації іншими популяціями на певному трофічному щаблі.
Часто доплив і відплив з популяції зображають у вигляді водяного стовпа з різними варіантами вхідного і вихідного отворів (рис. 3.). Надходження води до резервуару (р) ілюструє продукцію популяції, а розхід (е) – елімінацію (втрати, відпад, винесення особин за межі популяції). Висота водяного стовпа (h) ілюструє щільність популяції. В такому випадку втрати e є функцією продукції і щільності популяції: е = f (p,h). Коли потік є сталий, то ширина вхідного і вихідного отворів може бути чи великою, чи малою, а пропорція водяних стовпів залишається однаковою h1 = h2. Цей випадок добре ілюструють наведені на початку розділу варіанти зміни чисельності популяції при загальному її збільшенні на 10 особин. В іншому випадку врівноважений стан забезпечується більшим допливом і меншим відпливом біомаси.
Як бачимо, цей багатокомпонентний і динамічний процес тяжко вкласти в рамки простого рівняння, яке краще всього могло б розв'язуватися в умовах, коли можна було б припинити відпад і провести обміри біомаси. Однак у природі (як і в лабораторії) цей динамічний процес зупинити не можна, як не можна зупинити життя.
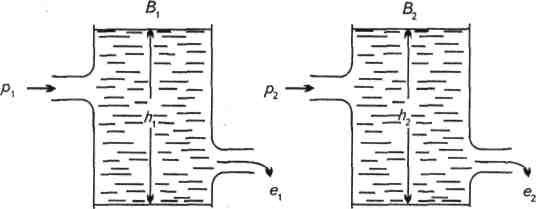
Рис.3. Врівноваження взаємного стану біомаси (В), продукції (р) і відпаду (е).
Цей стан біомаси може утримуватися як при високому рівні продукції системи, так і при низькому.
У звичайних умовах загальну продукцію популяції можна зобразити приростом біомаси (ΔВ) і елімінацією або відпадом (Е):
![]()
Зауважимо, що найвищий рівень продуктивності популяції у випадку найменшого відпаду. В природі трапляються такі випадки в агроценозах, коли кількість організмів експлуатуючої популяції усувається методами хімічного захисту, забезпечуючи можливість максимального приросту експлуатованої популяції.
В рівнянні Р.Вігерта, Ф.Еванса (1964) оцінюється первинна продукція з урахуванням впливу, який на неї має явище детрифікації рослинного матеріалу:
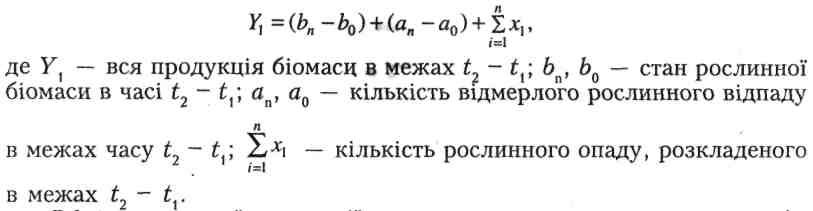
Обмір первинної продукції природних угруповань охоплює, крім оцінки станів біомаси, кількість усього матеріалу та з'ясування темпів розкладу мертвого матеріалу.
В популяціях, які в річному циклі дають лише одне покоління, а межі розроджуваності є короткі, оцінка продукції може бути проведена в межах початкового виходу особин (N0) та їх наступної редукції. Прикладом такого виду є бурякова стонка, яка проходить чотири личинкові стадії, під час яких живе на листях буряка, годуючись ними. Стадія лялечки перебуває в ґрунті, а доросла особина знову живе на листях буряка аж до настання холодів, після чого заривається в землю на зимування. Оцінка повної чисельності особин у популяції можлива з урахуванням відкладених яєць. Оцінка росту чисельності у різних стадіях личинки можлива на підставі кривої редукції (рис.4). Продукція, створена в окремих стадіях личинки, є функцією термічних умов середовища. Загальна продукція популяції в стадії личинки (Р) дорівнює
![]()
де ΔВ – приріст біомаси на даній стадії личинки; N0, N1 – початкова і кінцева чисельність особин у даному стані личинки; L1, L4 – чергові стадії личинок.
Загальна продукція, яка обіймає повний цикл розвитку виду, дає достатньо даних про приріст біомаси і стан енергії, закладеної в літньому жуку, який з'являється в серпні й інтенсивно жирує на посадках буряка до викопування або до повного з'їдання листя.
У популяціях, в яких декілька поколінь виступає одночленно, а розвиток має тривалий характер, основним способом визначення продукції є дослідження обороту особинами.
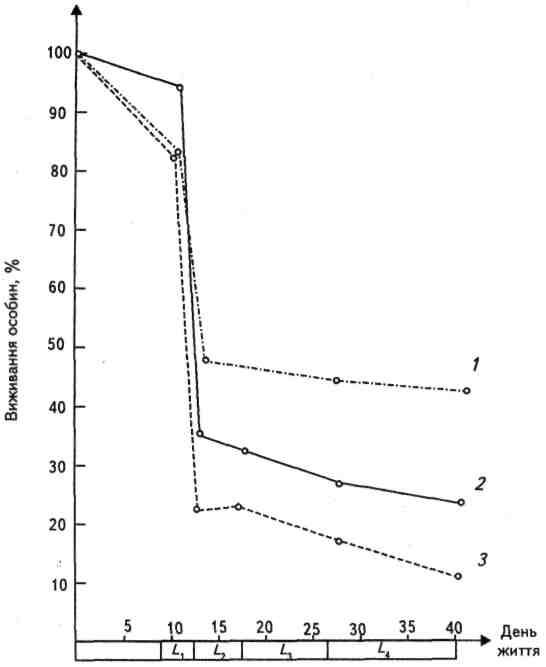
Рис. 4. Хід природного розмноження бурякової стонки на посівах буряка в західній частині Польщі: 1 – в кінці червня, 2 – в липні, 3 – на початку серпня, L1-L4 – чергова стадія личинки.
Оборот, як і інші параметри продуктивності (Р, A, R, С, FU), розглядають у певних часових межах (звичайно за рік, за сезон). Звідси витікає загальне визначення продукції: це повна кількість органічної матерії, створена в даному (певному) часі, не використана на распірацію (P=A-R), незалежно від того, чи залишилася ця матерія в популяції на кінець даного часу (Р = дельтаВ + Е), чи буде винесена.
Наведемо п'ять різних варіантів математичного виразу продукції за К. Петрусевичем:
1) Р = А - R (сальдо нетто між допливом енергії і коштом на її утримання);
2) Р = С - R - FU ( сальдо нетто вжитого корму і фекалій разом з уринами (сечею) та кошту утримання);
3) Р = ΔВ + Е (приріст біомаси разом з відпадом);
4) Р = Pr + Pg (продукція зростає внаслідок росту тіл і розроджуваності);
5) Р = B·Q (середня біомаса в обороті).
Отже, біомаса, створена популяцією за одиницю часу, є продукція нетто цієї популяції. На цю продукцію накладаються два процеси: приріст чисельності особин, випродукованих популяцією, а також приріст біомаси цих особин [1].