

2. Первичная и вторичная структура белка
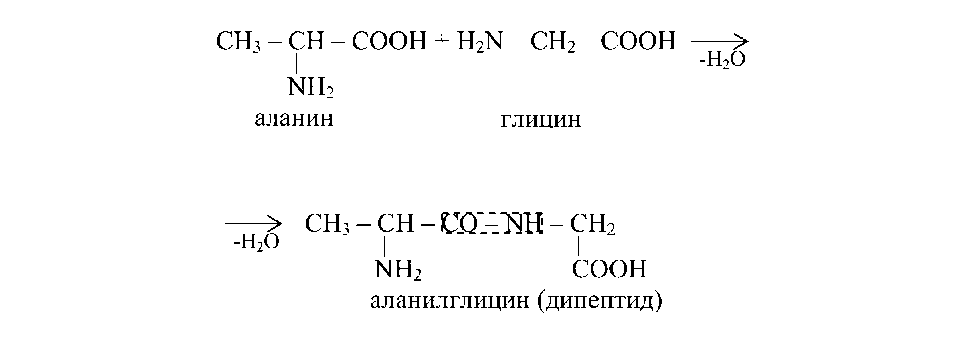
Первичная структура белка — последовательность расположения аминокислотных остатков в полипептиде. Для определения первичной структуры белка с помощью гидролиза выясняют количественное соотношение аминокислот в молекуле белка. Затем определяют химическую природу концевых аминокислот полипептидной цепи, которая содержит по одной свободной NH2- и COOH-группе. Для определения N-концевой аминокислоты используются метод Сэнджера и фенилгидантоиновый метод, для определения С-концевой аминокислоты — метод Акабори.
Первичная структура белка характеризуется рядом особенностей.
Первичная структура белка генетически детерминирована и уникальна, замена аминокислот внутри полипептидной цепи приводит к изменению структуры и функции белка.
Первичная структура белка стабильна, что обеспечивается дипептидными и в меньшей степени дисульфидными связями.
Число комбинаций аминокислот в полипептиде очень велико, повторяющиеся последовательности аминокислот редки. Идентичные последовательности аминокислот могут встречаться в области активных центров ферментов.
Первичная структура белка детерминирует вторичную, третичную и четвертичную структуру белковой молекулы.
Вторичная структура белка — конфигурация полипептидной цепи, более компактная ее упаковка в спиральную или какую-либо другую конформацию. Процесс конфигурации идет в соответствии с программой, заложенной в первичной структуре белка. Существует две основных конфигурации полипептидной цепи: α-спираль и β-сладчатый слой.
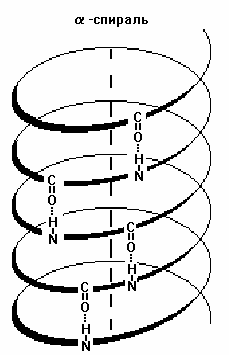
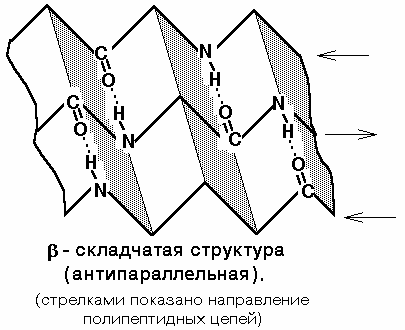
Действующим началом образования α- и β-структуры является способность аминокислот образовывать водородные связи. Стабильность вторичной структуры обеспечивается в основном водородными и в меньшей степени пептидными и дисульфидными связями.
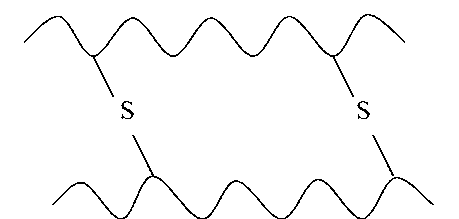
Водородная связь — слабое электростатическое взаимодействие между электроотрицательным атомом (O или N) и атомом водорода, ковалентно связанным с другим электроотрицательным атомом. Основные типы водородных связей:
между ОН-группой серина и пептидной связью
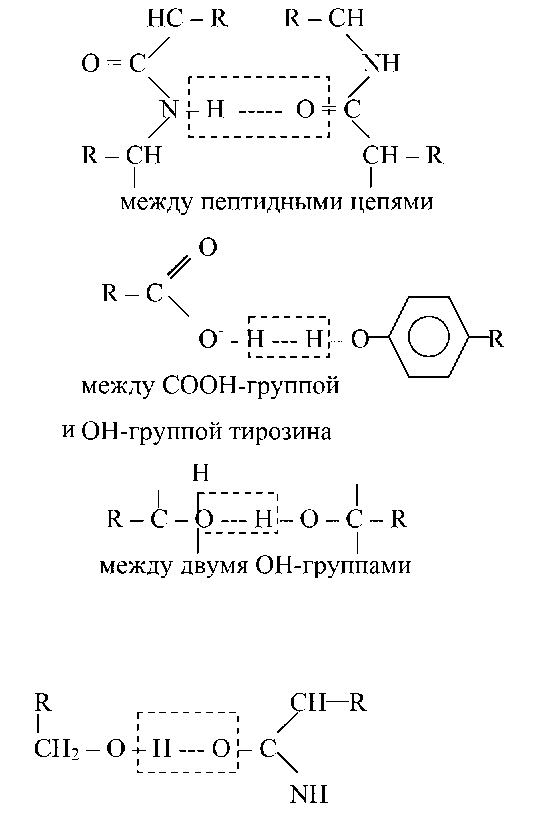
Структуре α-спирали присущ ряд закономерностей. На каждый шаг спирали приходится 3,6 аминокислотных остатка, шаг спирали равен 0,54 нм на виток и 0,15 нм на один аминокислотный остаток. Угол подъема спирали 26°, через каждые пять витков повторяется структурная конформация полипептида. При образовании β-структуры две или более линейные полипептидные цепи, расположенные параллельно либо антипараллельно, связываются между собой водородными связями между пептидными связями. В природе встре-чаются белки, вторичная структура которых не является ни α-, ни β-структурой (например, коллаген). В настоящее время доказано существование двух промежуточных уровней организации белковой молекулы между вторичной и третичной структурой — надвторичные структуры и структурные домены.
3. Третичная и четвертичная структура белка
Третичная структура белка — это пространственная ориентация полипептидной спирали, следующий за вторичной структурой способ компактизации белковой молекулы. Процесс укладки полипептидной цепи — фолдинг.
Пространственная структура белков зависит от ионной силы и pH раствора, температуры и иных факторов. В стабилизации пространственной структуры белков наряду с ковалентными связями (пептидными и дисульфидными) принимают участие так называемые нековалентные связи, к которым относятся водородные связи, электростатические взаимодействия заряженных групп, вандерваальсовы силы, взаимодействия неполярных боковых радикалов аминокислот, гидрофильно-гидрофобные взаимодействия и др. При этом нековалентные связи играют основную роль. Третичная структура белка формируется самопроизвольно и полностью детерминируется первичной структурой белка. Основным движущим моментом в возникновении трехмерной структуры является взаимодействие радикалов аминокислот с молекулами воды, состоящее в том, что гидрофобные радикалы аминокислот ориентируются внутрь молекулы белка, в то время как гидрофильные радикалы ориентируются наружу. В результате образуется термодинамически наиболее выгодная конформация молекулы белка, характеризующаяся наименьшей свободной энергией. Конформации белков стабильны, основные формы конформаций — T-форма (tensed — «напряженная») и R-форма (relaxed — «расслабленная»). Трехмерная структура белка содержит функциональную информацию, определяющую все биологические свойства белков. Нарушение третичной структуры белка влечет за собой потерю его биологических свойств.
Четвертичная структура белка — пространственная ориентация нескольких полипептидных цепей, обладающих собственной первичной, вторичной или третичной структурой, с образованием макромолекулярного образования.
Отдельные полипептидные цепи — протомеры (мономеры, субъединицы) — не обладают биологической активностью и приобретают ее при определенном способе пространственного объединения. Образовавшаяся молекула является олигомером (мультимером).
Четвертичная структура стабилизируется за счет нековалентных связей между контактными площадками протомеров, комплементарных друг другу. Четвертичная структура обнаружена у нескольких сотен белков. Молекула гемоглобина представляет собой тетрамер — состоит из двух α- и двух β-цепей. Фермент фосфорилаза a состоит из двух идентичных субъединиц по две пептидные цепи. Фермент лактатдегидрогеназа содержит два типа полипептидных цепей — M (muscle — «мышца») и B (brain — «мозг») и может существовать в пяти формах, называемых изоферментами, или множественными формами ферментов.