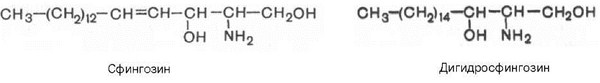Лекция №39-40 глицериды фосфолипиды
1. Глицериды Глицериды (ацилглицерины, или ацилглицеролы) представляют собой сложные эфиры трехатомного спирта глицерина и высших жирных кислот. Если жирными кислотами этерифицированы все три гидроксильные группы глицерина (ацильные радикалы R1, R2 и R3 могут быть одинаковы или различны), то такое соединение называют триглицеридом (триацилглице-рол), если две—диглицеридом (диацилглицерол) и, наконец, если этери-фицирована одна группа — моноглицеридом (моноацилглицерол):
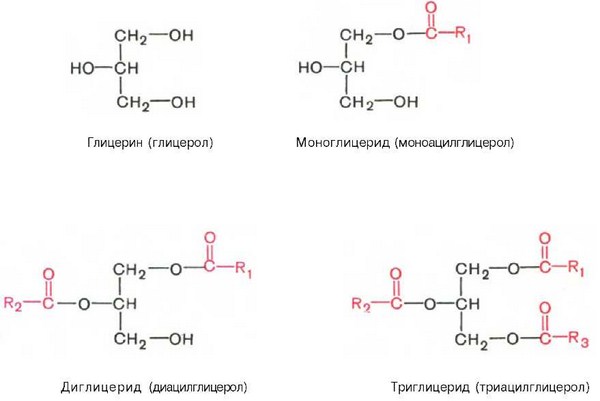
Наиболее распространенными являются триглицериды, часто называемые нейтральными жирами или просто жирами. Нейтральные жиры находятся в организме либо в форме протоплазматического жира, являющегося структурным компонентом клеток, либо в форме запасного, резервного, жира. Роль этих двух форм жира в организме неодинакова. Протоплазматический жир имеет постоянный химический состав и содержится в тканях в определенном количестве, не изменяющемся даже при патологическом ожирении, в то время как количество резервного жира подвергается большим колебаниям.
Как отмечалось, основную массу природных нейтральных жиров составляют триглицериды. Жирные кислоты в триглицеридах могут быть насыщенными и ненасыщенными. Из жирных кислот чаще встречаются пальмитиновая, стеариновая и олеиновые кислоты. Если все три кислотных радикала принадлежат одной и той же жирной кислоте, то такие три-глицериды называют простыми (например, трипальмитин, тристеарин, триолеин и т. д.), если разным жирным кислотам, то смешанными. Названия смешанных триглицеридов образуются в зависимости от входящих в их состав жирных кислот, при этом цифры 1, 2 и 3 указывают на связь остатка жирной кислоты с соответствующей спиртовой группой в молекуле глицерина (например, 1-олео-2-пальмитостеарин).
Жирные кислоты, входящие в состав триглицеридов, практически определяют их физико-химические свойства. Так, температура плавления триглицеридов повышается с увеличением числа и длины остатков насыщенных жирных кислот. Напротив, чем выше содержание ненасыщенных жирных кислот, или кислот с короткой цепью, тем ниже точка плавления. Животные жиры (сало) обычно содержат значительное количество насыщенных жирных кислот (пальмитиновой, стеариновой и др.). благодаря чему при комнатной температуре они твердые. Жиры, в состав которых входит много ненасыщенных кислот, при обычной температуре жидкие и называются маслами. Так, в конопляном масле 95% всех жирных кислот приходится на долю олеиновой, линолевой и линоленовой кислот и только 5% — на долю стеариновой и пальмитиновой кислот. В жире человека, плавящемся при температуре 15°С (при температуре тела он жидкий), содержится 70% олеиновой кислоты.
Глицериды способны вступать во все химические реакции, свойственные сложным эфирам. Наибольшее значение имеет реакция омыления, в результате которой из триглицеридов образуются глицерол и жирные кислоты. Омыление жира может происходить как при ферментативном гидролизе, так и при действии кислот или
2. Фосфолипиды Фосфолипиды представляют собой сложные эфиры многоатомных спиртов глицерина или сфингозина с высшими жирными кислотами и фосфорной кислотой. В состав фосфолипидов входят также азотсодержащие соединения: холин, этаноламин или серин. В зависимости от того, какой многоатомный спирт участвует в образовании фосфолипида (глицерин или сфингозин), последние делят на 2 группы: глицерофосфолипиды и сфинго-фосфолипиды. Необходимо отметить, что в глицерофосфолипидах либо холин, либо этаноламин или серин соединены эфирной связью с остатком фосфорной кислоты; в составе сфинголипидов обнаружен только холин. Наиболее распространенными в тканях животных являются глицерофосфо-липиды. Глицерофосфолипиды
Глицерофосфолипиды являются производными фосфатидной кислоты. В их состав входят глицерин, жирные кислоты, фосфорная кислота и обычно азотсодержащие соединения. Общая формула глицерофосфолипидов выглядит так:
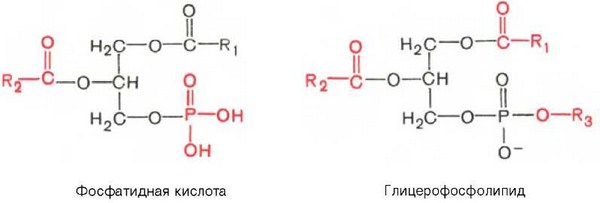
В этих формулах R1 и R2 — радикалы высших жирных кислот, a R3—чаще радикал азотистого соединения. Для всех глицерофосфолипидов характерно, что одна часть их молекул (радикалы R1 и R2) обнаруживает резко выраженную гидрофобность, тогда как другая часть гидрофильна благодаря отрицательному заряду фосфорной кислоты и положительному заряду радикала R3.
Из всех липидов глицерофосфолипиды обладают наиболее выраженными полярными свойствами. При помещении глицерофосфолипидов в воду в истинный раствор переходит лишь небольшая их часть, основная же масса липидов находится в виде мицелл. Существует несколько групп (подклассов) глицерофосфолипидов. В зависимости от характера азотистого основания, присоединенного к фосфорной кислоте, Глицерофосфолипиды подразделяют на фосфатидилхолины (лецитины), фосфатидилэтаноламины (кефалины) и фосфатидилсерины. В состав некоторых глицерофосфолипидов вместо азотсодержащих соединений входит не содержащий азота шестиуглеродный циклический спирт инозит, называемый также инозитолом. Эти липиды называются фосфатидилинозитолами.
Фосфатидилхолины (лецитины). В отличие от триглицеридов в молекуле фосфатидилхолина одна из трех гидроксильных групп глицерина связана не с жирной, а с фосфорной кислотой. Кроме того, фосфорная кислота в свою очередь соединена эфирной связью с азотистым основанием — холином [HO-CH2-CH2-N+(CH3)3]. Таким образом, в молекуле фосфатидил-холина соединены глицерин, высшие жирные кислоты, фосфорная кислота и холин:
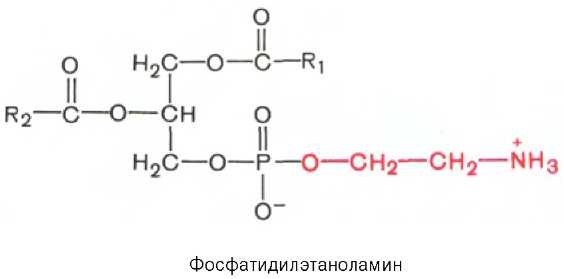
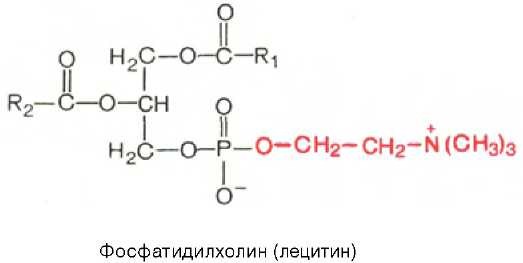
Фосфатидилэтаноламины. Основное различие между фосфатидилхоли-нами и фосфатидилэтаноламинами — наличие в составе последних азотистого основания этаноламина (HO—CH2—CH2—N+H3):Из глицерофосфолипидов в организме животных и высших растений в наибольшем количестве встречаются фосфатидилхолины и фосфатидил-этаноламины. Эти 2 группы глицерофосфолипидов метаболически связаны друг с другом и являются главными липидными компонентами мембран клеток.
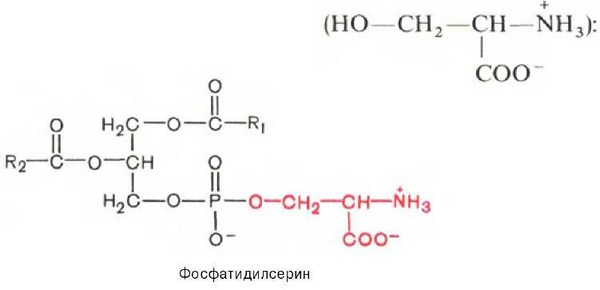
Фосфатидилсерины. В молекуле фосфатидилсерина азотистым соединением служит остаток аминокислоты серина
Фосфатидилсерины распространены гораздо менее широко, чем фос-фатидилхолины и фосфоэтаноламины, и их значение определяется в основном тем, что они участвуют в синтезе фосфатидилэтаноламинов.
Фосфатидилинозитолы. Эти липиды относятся к группе производных фосфатидной кислоты, но не содержат азота. Радикалом (R3) в этом подклассе глицерофосфолипидов является шестиуглеродный циклический спирт инозитол:
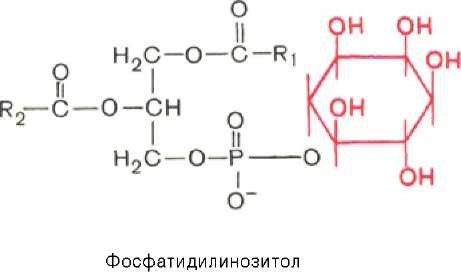
Фосфатидилинозитолы довольно широко распространены в природе. Они обнаружены у животных, растений и микроорганизмов. В животном организме найдены в мозге, печени и легких.
Плазмалогены. От рассмотренных глицеролипидов плазмалогены отличаются тем, что вместо одного остатка высшей жирной кислоты содержат остаток а, Р-ненасыщенного спирта, который образует простую связь (в отличие от сложноэфирной связи, образуемой остатком жирной кислоты) с гидроксильной группой глицерина в положении С-1:
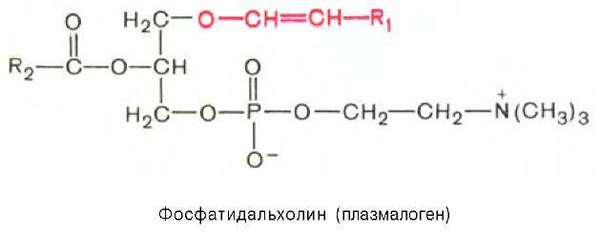
Основными подклассами плазмалогенов являются фосфатидальхолины, фосфатидальэтаноламины и фосфатидальсерины. В разбавленных кислотах плазмалогены гидролизуются с образованием альдегида соответствующего а, Р-ненасыщенного спирта, т. е. при кислотном гидролизе плазмалогенов образуются «жирные» альдегиды, называемые плазмалями, что и легло в основу термина «плазмалоген».
Кардиолипин. Своеобразным представителем глицерофосфолипидов является кардиолипин, впервые выделенный из сердечной мышцы. По своей химической структуре кардиолипин можно рассматривать как соединение, в котором 2 молекулы фосфатидной кислоты связаны с помощью одной молекулы глицерина. В отличие от остальных глицерофосфолипидов кар-диолипин является как бы «двойным» глицерофосфолипидом. Кардиолипин локализован во внутренней мембране митохондрий. Функция его пока неясна, хотя известно, что в отличие от других фосфолипидов кардиолипин обладает иммунными свойствами.
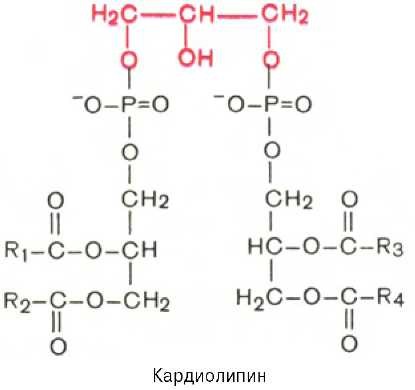
В этой формуле R1, R2, R3, R4 – радикалы высших жирных кислот.
Необходимо отметить, что в природе встречается свободная фосфатид-ная кислота, но в относительно небольших количествах по сравнению с глицерофосфолипидами. Среди жирных кислот, входящих в состав гли-церофосфолипидов, обнаружены как насыщенные, так и ненасыщенные (чаще стеариновая, пальмитиновая, олеиновая и линолевая).
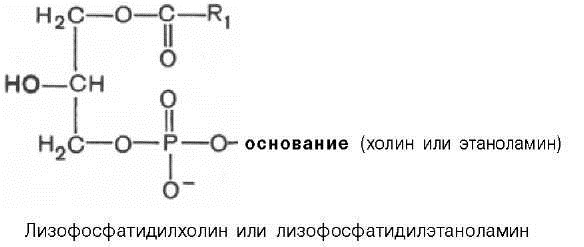
Установлено также, что большинство фосфатидилхолинов и фосфати-дилэтаноламинов содержат одну насыщенную высшую жирную кислоту в положении С-1 и одну ненасыщенную высшую жирную кислоту в положении С-2. Гидролиз фосфатидилхолинов и фосфатидилэтаноламиновпри участии особых ферментов (эти ферменты относятся к фосфолипазам A2), содержащихся, например, в яде кобры, приводит к отщеплению ненасыщенной жирной кислоты и образованию лизофосфолипидов — лизофосфатидилхолинов, или лизофосфатидилэтаноламинов, оказывающих сильное гемолитическое действие:
Сфинголипиды (сфингофосфолипиды)
Сфингомиелины. Это наиболее распространенные сфинголипиды. В основном они находятся в мембранах животных и растительных клеток. Особенно богата ими нервная ткань. Сфингомиелины обнаружены также в ткани почек, печени и других органов. При гидролизе сфингомиелины образуют одну молекулу жирной кислоты, одну молекулу двухатомного ненасыщенного спирта сфингозина, одну молекулу азотистого основания (чаще это холин) и одну молекулу фосфорной кислоты. Общую формулу сфингомиелинов можно представить так:
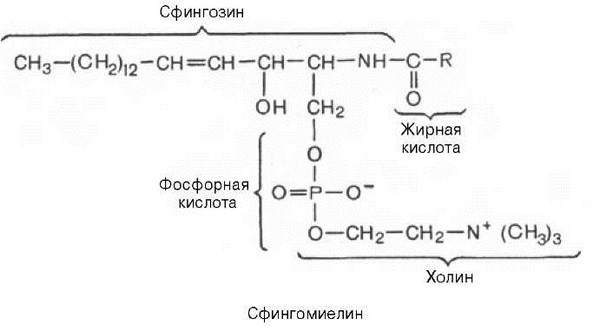
Общий план построения молекулы сфингомиелина в определенном отношении напоминает строение глицерофосфолипидов. Молекула сфин-гомиелина содержит как бы полярную «головку», которая несет одновременно и положительный (остаток холина), и отрицательный (остаток фосфорной кислоты) заряды, и два неполярных «хвоста» (длинная алифатическая цепь сфингозина и ацильный радикал жирной кислоты). В некоторых сфингомиелинах, например выделенных из мозга и селезенки, вместо сфингозина найден спирт дигидросфингозин (восстановленный сфингозин):